|
4.3.6 –°љб
љбЇѕ∆дЋы—–Њњ’яµƒєџµгЇЌќ“√«µƒ µ—йљбєы£ђ„џЇѕњЉ¬«≥ц√з¬ °Ґ≥ц√з ±Љд°Ґ≥ц√зЋў¬ °ҐЈµ«аіжїоЇЌ…ъќпЅњµ»ЉЄЄц÷Є±к£ђќ“√«»ѕќ™“∞…ъЇЌ‘‘≈а—т≤Ёµƒ Їѕ≤•÷÷…оґ»ќ™1-2cm£ђ„о…о≤ї“Ћ≥ђєэ4cm°£’в”л“‘«∞—–ЊњЋщљ®“йµƒ—т≤Ё Їѕ≤•÷÷…оґ»4-5cm≤ї“ї÷¬°£“∞…ъ—т≤ЁµƒњєЇµ–‘«њ”Џ‘‘≈а—т≤Ё£ђЈµ«аіжїоЇЌ…ъќпЅњ”≈”Џ‘‘≈а—т≤Ё°£“тіЋ£ђ‘Џљш––÷≤±її÷Єіє№јн µЉщ ±£ђ—°‘сЇѕ µƒ≤Ё÷÷ЇЌ µ±µƒ≤•÷÷…оґ» «Ј«≥£±Ў“™µƒ°£
4.4 њЁ≤Ё≤гґ‘—т≤ЁЇЌќё√Ґ»Є¬у÷÷√з≥ц√зЇЌ‘з∆Џ…ъ≥§µƒ”∞ѕм
‘—й”Џ2006ƒк5-10‘¬‘ЏЇ”±± °’≈Љ“њЏ –»ы±±є№јн«ш÷–єъ≈©“µіу—І≤ЁµЎ—–Њњ µ—й’Њ°™°™єъЉ“≤ЁµЎ…ъћђѕµЌ≥“∞Ќвњ∆—І—–Њњєџ≤в’Њ£®»ы±±£©љш––£ђ ‘—йЈ÷ќ™ћпЉдЇЌќ¬ “Ѕљ≤њЈ÷°£ “Ќв ‘—й‘Џћм»ї≤Ё≥°£ђ “ƒЏ ‘—й‘Џ µ—й’Њќ¬ “љш––°£Ѕљ÷÷њЁ≤Ё≤г≤ƒЅѕЈ÷±рќ™і”µ±µЎћм»ї≤ЁµЎ ’ЉѓµƒњЁ¬дќп£®ћм»їњЁ≤Ё£©“‘Љ∞»Ћє§Ћ№Ѕѕƒ£ƒвњЁ≤Ё≤г£®ƒ£ƒвњЁ≤Ё£©£ђќё√Ґ»Є¬у÷÷„”Љ∞“∞…ъ—т≤Ё÷÷„”ќ™2004ƒк‘Џµ±µЎћм»ї≤ЁµЎ ’Љѓµƒ÷÷„”°£ ‘—й…и2÷÷≤їЌђµƒљљЋЃЅњ°Ґ 4÷÷≤їЌђµƒњЁ≤Ё≤г÷ЎЅњ°Ґ3÷÷≤•÷÷ќї÷√ЇЌ2÷÷њЁ≤Ё≤гја–Ќ°£
4.4.1 њЁ≤Ё≤гја–Ќґ‘÷÷√зљ®÷≤µƒ”∞ѕм
ќ¬ “ µ—й±н√ч£®±н1-21ЇЌ±н1-22£©£ђњЁ≤Ё≤гја–Ќґ‘÷÷√з≥ц√зЇЌ…ъ≥§Ћўґ»”–ѕ‘÷ш”∞ѕм°£ћм»їњЁ≤Ё”лƒ£ƒвњЁ≤Ёѕа±»љµµЌЅЋ—т≤ЁЇЌќё√Ґ»Є¬уµƒ÷÷√з≥ц√з¬ ЇЌ…ъ≥§Ћўґ»°£іЋЌв£ђћм»їњЁ≤Ё“≤ѕ‘÷шљµµЌЅЋ—т≤Ёµƒіжїо¬ ЇЌµЎѕ¬…ъќпЅњ°£
4.2 љљЋЃЅњґ‘÷÷√зљ®÷≤µƒ”∞ѕм
љљЋЃЅњґ‘≥ц√з¬ °Ґ…ъќпЅњЇЌ…ъ≥§Ћўґ»”–ѕ‘÷ш”∞ѕм°£„№µƒјіљ≤£ђЄяєаЋЃЅњ‘цЉ”ЅЋќё√Ґ»Є¬уЇЌ—т≤Ёµƒ≥ц√з¬ ЇЌ…ъќпЅњ°£љљЋЃЅњіуµƒ÷÷√з≥ц√зЋўґ»њм”ЏљљЋЃЅњ–°µƒ£®Љы±н1-21ЇЌ±н1-22£©°£
4.3 њЁ≤Ё≤г÷ЎЅњґ‘÷÷√зљ®÷≤µƒ”∞ѕм
ќ¬ “ ‘—й±н√ч£®±н1-21ЇЌ±н1-22£©£ђњЁ≤Ё≤гµƒ÷ЎЅњѕ‘÷ш”∞ѕм—т≤Ёµƒ…ъ≥§Ћўґ»ЇЌµЎѕ¬…ъќпЅњ£ђґшґ‘ќё√Ґ»Є¬уµƒЄчѕо÷Є±кЊщќёѕ‘÷ш”∞ѕм°£÷÷√з…ъ≥§Ћўґ»ЇЌ„№…ъќпЅњЋж„≈њЁ≤Ё≤г÷ЎЅњµƒ‘цЉ”ґш‘цЉ”°£“∞ЌвћхЉюѕ¬£®±н1-23ЇЌ±н1-24£©£ђњЁ≤Ё≤г÷ЎЅњґ‘—т≤Ёµƒ≥ц√з¬ ЇЌіжїо¬ ґЉ”–ѕ‘÷ш”∞ѕм£ђґ‘ќё√Ґ»Є¬уµƒ≥ц√з¬ ЇЌіжїо¬ √ї”–ѕ‘÷ш”∞ѕм°£—т≤Ёµƒ≥ц√з¬ ЋжњЁ≤Ё≤г÷ЎЅњµƒ‘цЉ”ґшљµµЌ£ђµЂіжїо¬ »іЋжњЁ≤Ё≤г÷ЎЅњµƒ‘цЉ”ґш‘цЉ”£ђњ…ƒ№ «”…”ЏњЁ≤Ё≤г”–јы”Џ±£≥÷ЌЅ»јЋЃЈ÷µƒ‘µє °£
4.4 ÷÷„”ќї÷√ґ‘÷÷√зљ®÷≤µƒ”∞ѕм
±н1-21°Ґ22ЇЌ23ЇЌ24ѕ‘ Њ£ђ÷÷„”ќї÷√ґ‘ќ¬ “ЇЌ“∞Ќвµƒ—т≤ЁЇЌќё√Ґ»Є¬уµƒ≥ц√з¬ ґЉ”–ѕ‘÷ш”∞ѕм°£≤•”ЏЌЅ»ј÷–µƒ÷÷„”≥ц√з¬ ‘ґ‘ґЄя”Џ∆дЋьЅљ÷÷≤•÷÷Јљ љ£ђґш≤•”ЏњЁ≤Ё≤г”лЌЅ»ј±н√ж÷ЃЉдµƒ÷÷„”≥ц√з¬ “≤іу”Џ≤•”ЏњЁ≤Ё≤г±н√жµƒ≥ц√з¬ °£÷÷„”ќї÷√ґ‘—т≤Ё÷÷„”≥ц√зЇЌ…ъ≥§µƒ”∞ѕм±»ґ‘ќё√Ґ»Є¬уµƒ”∞ѕм“™іу°£“∞ЌвћхЉюѕ¬µƒЌЅ»ј÷–µƒ—т≤Ё≥ц√з¬ «÷÷„”ќї”ЏњЁ≤Ё≤г±н√жµƒ2-3±ґґа£ђќё√Ґ»Є¬уµƒ≥ц√з¬ ÷їѕа≤о10%„у”“°£÷÷„”ќї÷√ґ‘ќ¬ “—т≤ЁЇЌќё√Ґ»Є¬уµƒ…ъ≥§Ћўґ»°ҐµЎ…ѕ°ҐµЎѕ¬ЇЌ„№…ъќпЅњґЉ”–ѕ‘÷ш”∞ѕм£ђ÷÷„”≤•”ЏЌЅ»ј÷–µƒ э÷µ„оЄя°£
±н1-21 њЁ≤Ё≤гја–Ќ°ҐљљЋЃЅњ°ҐњЁ≤Ё≤г÷ЎЅњ”л÷÷„”ќї÷√ґ‘ќё√Ґ»Є¬у≥ц√зЇЌіжїоµ»µƒ”∞ѕм£®ќ¬ “£©
|
і¶јн |
ЋЃ∆љ |
≥ц√з¬ £®%£© |
іжїо¬ £®%£© |
…ъ≥§Ћўґ»£®cm£© |
µЎ…ѕ…ъќпЅњ£®g£© |
µЎѕ¬…ъќпЅњ£®g£© |
„№…ъќпЅњ£®g£© |
|
њЁ≤Ё≤гја–Ќ |
ћм»їњЁ≤Ё |
68.22b |
96.4a |
14.33b |
3.30a |
6.06a |
9.36a |
|
Ћ№ЅѕњЁ≤Ё |
81.22a |
96.5a |
16.74a |
2.97a |
5.78a |
8.75a |
|
љљЋЃЅњ |
µЌ |
72.67b |
96.08a |
16.46b |
2.94b |
6.05a |
8.99a |
|
Єя |
76.78a |
96.76a |
14.61a |
3.34a |
5.79a |
9.13a |
|
њЁ≤Ё≤г
÷ЎЅњ |
0 |
75.72a |
96.67a |
15.36a |
2.93a |
5.56a |
8.49a |
|
1/2∆љЊщ÷µ |
75.56a |
97.94a |
16.72a |
3.26a |
5.99a |
9.26a |
|
∆љЊщ÷µ |
70.89a |
94.78a |
14.72a |
3.13a |
5.44a |
8.57a |
|
2±ґ∆љЊщ÷µ |
76.72a |
96.31a |
15.35a |
3.23a |
6.68a |
9.91a |
|
÷÷„”ќї÷√ |
њЁ≤Ё≤г±н√ж |
60.88c |
98.31b |
14.36b |
2.63b |
4.79b |
7.42b |
|
њЁ≤Ё≤г”лЌЅ»јЉд |
71.96b |
96.60ab |
14.68b |
2.90b |
5.05b |
7.95b |
|
ЌЅ»ј÷– |
91.33a |
94.35a |
17.56a |
3.88a |
7.91a |
11.80a |
„Ґ£Їµ±µЎ≤Ё‘≠њЁ≤Ё≤г÷ЎЅњ∆љЊщ÷µќ™£Ї100g/m2£їљљЋЃЅњЈ÷±рќ™µ±µЎљµ”кЅњ„о–°÷µ”л„оіу÷µЉі50mm/‘¬ЇЌ100mm/‘¬
±н1-22 њЁ≤Ё≤гја–Ќ°ҐљљЋЃЅњ°ҐњЁ≤Ё≤г÷ЎЅњ”л÷÷„”ќї÷√ґ‘“∞…ъ—т≤Ё≥ц√зЇЌіжїоµ»µƒ”∞ѕм£®ќ¬ “£©
|
і¶јн |
ЋЃ∆љ |
≥ц√з¬ £®%£© |
іжїо¬ £®%£© |
…ъ≥§Ћўґ»£®cm£© |
µЎ…ѕ…ъќпЅњ£®g£© |
µЎѕ¬…ъќпЅњ£®g£© |
„№…ъќпЅњ£®g£© |
|
њЁ≤Ё≤г
ја–Ќ |
ћм»їњЁ≤Ё |
44.61b |
86.97b |
11.57b |
1.90a |
1.98b |
3.89a |
|
Ћ№ЅѕњЁ≤Ё |
52.56a |
91.63a |
16.91a |
1.78a |
2.49a |
4.27a |
|
љљЋЃЅњ |
µЌ |
38.78b |
89.67a |
12.64b |
1.61b |
2.08b |
3.69b |
|
Єя |
58.39a |
88.93a |
15.84a |
2.08a |
2.39a |
4.47a |
|
њЁ≤Ё≤г
÷ЎЅњ |
0 |
50.39a |
76.14b |
12.41b |
1.66a |
1.74b |
3.40a |
|
1/2∆љЊщ÷µ |
46.33a |
93.11a |
14.13ab |
1.74a |
2.18ab |
3.92a |
|
∆љЊщ÷µ |
47.89a |
94.72a |
15.02ab |
1.91a |
2.56a |
4.47a |
|
2±ґ∆љЊщ÷µ |
49.72a |
93.22a |
15.40a |
2.07a |
2.47ab |
4.53a |
|
÷÷„”ќї÷√ |
њЁ≤Ё≤г
±н√ж |
28.50c |
89.67a |
11.42b |
1.28b |
1.44b |
2.72b |
|
њЁ≤Ё≤г
”лЌЅ»јЉд |
39.67b |
89.94a |
12.44b |
1.53b |
1.83b |
3.35b |
|
ЌЅ»ј÷– |
77.58a |
88.29a |
18.86a |
2.73a |
3.44a |
6.17a |
„Ґ£Їµ±µЎћм»ї≤ЁµЎњЁ≤Ё≤г÷ЎЅњ∆љЊщ÷µќ™£Ї100g/m2£їљљЋЃЅњЈ÷±рќ™µ±µЎљµ”кЅњ„о–°÷µ”л„оіу÷µЉі50mm/‘¬ЇЌ100mm/‘¬
±н1-23 ћм»їњЁ≤Ё≤г÷ЎЅњ”л÷÷„”ќї÷√ґ‘ќё√Ґ»Є¬у≥ц√зЇЌіжїоµƒ”∞ѕм£®ћпЉд£©
|
і¶јн |
ЋЃ∆љ |
≥ц√з¬ £®%£© |
іжїо¬ £®%£© |
|
њЁ≤Ё≤г÷ЎЅњ |
0 |
47.33a |
53.13a |
|
1/2∆љЊщ÷µ |
51.47a |
50.67a |
|
∆љЊщ÷µ |
53.47a |
47.27a |
|
2±ґ∆љЊщ÷µ |
50.53a |
50.80a |
|
÷÷„”ќї÷√ |
њЁ≤Ё≤г±н√ж |
41.80b |
50.85a |
|
њЁ≤Ё≤г”лЌЅ»јЉд |
41.20b |
52.15a |
|
ЌЅ»ј÷– |
50.70a |
48.40a |
„Ґ£Їµ±µЎћм»ї≤ЁµЎњЁ≤Ё≤г÷ЎЅњ∆љЊщ÷µќ™£Ї100g/m2
±н1-24 ћм»їњЁ≤Ё≤г÷ЎЅњ”л÷÷„”ќї÷√ґ‘“∞…ъ—т≤Ё≥ц√зЇЌіжїоµƒ”∞ѕм£®ћпЉд£©
|
і¶јн |
ЋЃ∆љ |
≥ц√з¬ £®%£© |
іжїо¬ £®%£© |
|
њЁ≤Ё≤г÷ЎЅњ |
0 |
30.80b |
18.87b |
|
1/2∆љЊщ÷µ |
13.60a |
16.73b |
|
∆љЊщ÷µ |
20.53ab |
31.47a |
|
2±ґ∆љЊщ÷µ |
19.47ab |
33.40a |
|
÷÷„”ќї÷√ |
њЁ≤Ё≤г±н√ж |
12.90b |
25.10a |
|
њЁ≤Ё≤г”лЌЅ»јЉд |
18.00b |
26.80a |
|
ЌЅ»ј÷– |
32.40a |
23.45a |
„Ґ£Їµ±µЎћм»ї≤ЁµЎњЁ≤Ё≤г÷ЎЅњ∆љЊщ÷µќ™£Ї100g/m2
4.5 –°љб
1£©°Ґ≤їЌђµƒє№јніл ©£®∞“µЎ°ҐЎ„Єо°Ґ ©Ј °Ґ≤•÷÷√№ґ»£©ґ‘≤є≤•ƒ№Јс≥…є¶”–ѕ‘÷шµƒ”∞ѕм£ђ÷±љ”»ц≤•°Ґґ‘÷≤±ї≤їљш––»≈ґѓ£ђЉі є ©”√Ј ЅѕЇЌ‘цЉ”≤•÷÷√№ґ»“≤Ї№ƒ— є≤є≤•µƒќп÷÷÷÷√з≥…є¶љ®÷≤°£∞“µЎ»≈ґѓњ…“‘Ќ®єэљµµЌ÷≤±їЄ«ґ»°Ґ‘цЉ”÷÷„””лЌЅ»јљ”і•°Ґ‘ц«њЅЋєв’’ЇЌЌЅ»ј±н≤гќ¬ґ»£ђі”ґш‘цЉ”÷÷√з≥ц√зЇЌ…ъ≥§±нѕ÷£ђ”–јы”Џ÷÷√з≥…є¶љ®÷≤°£
2£©°Ґ÷÷√з√№ґ»”л≤•÷÷√№ґ»≥ ’эѕаєЎєЎѕµ£ђ≤є≤• ± µ±Љ”іу≤•÷÷√№ґ» «±Ў“™µƒ°£
3£©°Ґ≤їЌђµƒќп÷÷∆д≤є≤•ґ®Њ”≥…є¶µƒЉЄ¬ “≤≤їЌђ£ђќё√Ґ»Є¬уµƒ÷÷√зљ®÷≤ƒ№Ѕ¶“™”≈”Џ—т≤Ё£ђ±нѕ÷ќ™≥ц√зњм°Ґ…ъ≥§њм£ђ≤їЄ≤ЌЅ£®ЌЅ»ј≤ї»≈ґѓ£©≥ц√з¬ “≤Єя”Џ—т≤Ё£ђ“тіЋќё√Ґ»Є¬у≤є≤•љ®÷≤µƒ–Ієы“™Ї√”Џ—т≤Ё°£
4£©°Ґ≤є≤•—т≤ЁЇЌќё√Ґ»Є¬уїб №µљјі„‘ѕ÷іж÷≤±їµƒ“÷÷∆„ч”√£ђЊяће±нѕ÷ќ™“÷÷∆≥ц√зЇЌѕё÷∆÷÷√з…ъ≥§£ђ”»∆д «јі„‘ѕ÷іж÷≤±їµƒµЎѕ¬ЊЇ’щ «ѕё÷∆÷÷√зљ®÷≤µƒ„о÷ч“™“тЋЎ°£
5£©°Ґїм≤•ƒЅ≤Ё∆д÷÷√зљ®÷≤єэ≥ћ÷–÷÷√зЉд“≤”–ѕаї•„ч”√£ђќё√Ґ»Є¬у÷÷√з≥ц√з‘з°Ґ…ъ≥§њм£ђґ‘—т≤Ё÷÷√з…ъ≥§Њя”–“÷÷∆„ч”√°£
6£©°Ґ≤•÷÷…оґ»ЇЌ÷÷„”јі‘іѕ‘÷ш”∞ѕм÷÷√з≥цЌЅЇЌ…ъќпЅњµ»…ъ≥§±нѕ÷£ђ≤їЄ≤ЌЅ“≤ѕ‘÷шљµµЌЅЋ÷÷√з≥цЌЅ≤Ґ—”≥§ЅЋ≥ц√з ±Љд°£‘Џљш––÷≤±її÷Єіє№јн µЉщ ±£ђ—°‘сЇѕ µƒ÷÷„”јі‘іЇЌњЎ÷∆≤•÷÷…оґ» «Ј«≥£±Ў“™µƒ°£
7£©°Ґћм»їњЁ≤Ёґ‘÷÷√зљ®÷≤„№µƒјіЋµ”–ЄЇ√ж”∞ѕм£ђ÷ч“™±нѕ÷ќ™љµµЌЅЋ÷÷„”≥ц√з¬ £ђЉіЌ®єэ„и÷є÷÷„””лЌЅ»јљ”і•ґшѕ‘÷шљµµЌ≥ц√з¬ °£ ‘—й±н√чќї”ЏЌЅ»ј÷–µƒ÷÷„”∆д≥ц√з¬ ѕ‘÷шЄя”ЏњЁ≤Ё≤г±н√ж°ҐњЁ≤Ё≤г”лЌЅ»ј±н√жµƒ≥ц√з¬ °£‘Џћм»ї≤Ё‘≠÷≤±її÷Єіє№јн µЉщ÷–£ђ≤…»° µ±µƒіл ©»•≥эњЁ≤Ё є÷÷„”¬д»лЌЅ»ј£ђ≤≈ƒ№±£÷§÷÷„””–љѕЄяµƒ≥ц√з¬ ЇЌ÷÷√з≥…є¶љ®÷≤°£
8£©°Ґµ•≤•ЇЌїм≤•µƒ µ—йґЉ±н√ч£ђќё√Ґ»Є¬уµƒ÷÷√зљ®÷≤ƒ№Ѕ¶ЇЌЊЇ’щЅ¶“™”≈”Џ—т≤Ё°£
≥х≤љљб¬џ»зѕ¬£Ї
”∞ѕм—т≤Ё≥ц√зµƒ÷ч“™“тЋЎ «÷÷„””лЌЅ»јµƒљ”і•≥ћґ»ЇЌ≤•÷÷…оґ»£ђЄ≤Є«њЁ≤Ё≤г”–јы”Џ“∞Ќв—т≤Ё÷÷√зіжїо°£
ґ‘ЌЅ»јљш––»≈ґѓ£ђ∞“µЎ’т—є“‘ є÷÷„””лЌЅ»ј≥дЈ÷љ”і•£®÷÷„”»лЌЅ1-2cm£© «ћбЄя÷÷√з”»∆д «—т≤Ё÷÷√з≥ц√з¬ µƒ±Ў“™ћхЉю£ђ“≤ «≤є≤•—т≤Ё≥…є¶ї÷Єі≤ЁµЎ÷≤±їµƒєЎЉь£їљµµЌјі„‘ѕ÷іж÷≤±їµƒЊЇ’щ£ђ”»∆д «µЎѕ¬µƒЊЇ’щ «±£÷§—т≤Ё°Ґќё√Ґ»Є¬у÷÷√зљ®÷≤≥…є¶µƒ±Ў“™ћхЉю°£
5°Ґјы”√Јљ љґ‘ћм»ї≤ЁµЎƒЅ≤Ё…ъ≥§”лЈ±÷≥µƒ”∞ѕм—–Њњ
‘ЏЇ”±±’≈Љ“њЏ∞”…ѕєЅ‘іƒЅ≥°—т≤Ё≤Ё‘≠…ѕ£ђ—°‘с÷≤±їЊщ‘»°ҐќҐµЎ–ќ≤о“мљѕ–°µƒЄо≤ЁµЎЇЌЈ≈ƒЅµЎ„чќ™—–Њњ—щµЎ°£“‘÷÷»Їќ™—–Њњґ‘ѕу£ђ”Џ2005ƒк6‘¬°Ђ2005ƒк10‘¬£ђґ‘ƒЅ≤ЁµƒЈ±÷≥ґ‘≤яЇЌ÷÷„””кљш––ЅЋ—–Њњ£ђЈ÷ќцЅЋ≤Ё‘≠ƒЅ≤Ё‘Џ÷÷»ЇЋЃ∆љ…ѕµƒЈ±÷≥єж¬…ЇЌ÷≤±ї—Ёћжєж¬…°£
5.1 Ј≈ƒЅµЎ”лЄо≤ЁµЎ÷≤±їµч≤й
Єо≤ЁµЎ”≈ ∆ґ»„оіуµƒ«∞ќе÷÷÷≤ќпЈ÷±рќ™£Ї—т≤Ё£®59.6£©°Ґ≤Џ“ю„”≤Ё£®9.51£©°ҐƒѕƒµЁп£®3.80£©°ҐјдЁп£®2.96£©°Ґ’л√©£®2.74£©£ђ∆д÷–—т≤Ё’Љ”–Њшґ‘”≈ ∆£їґшЈ≈ƒЅµЎ”≈ ∆ґ»„оіуµƒ«∞ќе÷÷÷≤ќпЈ÷±рќ™£Ї—т≤Ё£®34.4£©°Ґ≤Џ“ю„”≤Ё£®9.36£©°Ґ’л√©£®8.06£©°Ґ∆—єЂ”Ґ£®6.04£©°Ґ¬нЁю£®3.96£©°£”…”Џ—т≤Ёµƒ”≈ ∆ґ»љµµЌ£ђЈ≈ƒЅµЎ÷–µƒ≤Џ“ю„”≤Ё°ҐƒѕƒµЁп°Ґ’л√©°ҐѕЄЅ—“ґЁп°Ґ√…є≈Ёп°ҐґюЅ—ќЃЅк≤Ћ°ҐѕЄ“ґЊ¬°ҐЇ„ ≠°ҐЅљ∆№ё§µ»ќп÷÷µƒЄ«ґ»°Ґ√№ґ»ЇЌ”≈ ∆ґ»ґЉіу”ЏЄо≤ЁµЎЌђ÷÷÷≤ќпµƒЄ«ґ»°Ґ√№ґ»ЇЌ”≈ ∆ґ»°£
±н1-25 Ј≈ƒЅµЎ°ҐЄо≤ЁµЎ÷≤ќп÷÷µƒЄяґ»°ҐЄ«ґ»°Ґ√№ґ»Љ∞”≈ ∆ґ»µƒ±»љѕ
|
ќп÷÷
|
Єо≤ЁµЎ |
Ј≈ƒЅµЎ |
|
Єяґ» Є«ґ» √№ґ» ”≈ ∆ґ» |
Єяґ» Є«ґ» √№ґ» ”≈ ∆ґ» |
|
—т≤Ё |
Leymus chinesis Tzvel. |
22.2 |
81 |
806 |
59.6 |
6.46 |
43 |
1244 |
34.4 |
|
≤Џ“ю„”≤Ё |
Poa annua L. |
7.69 |
26.3 |
47 |
9.51 |
4.15 |
28 |
59 |
9.36 |
|
’л√© |
Stipa grandis P. |
11.1 |
4 |
5.9 |
2.74 |
7.45 |
17 |
66.7 |
8.06 |
|
ћ¶≤Ё |
Carex cranaocarpa Mey. |
|
|
|
|
4.56 |
4.33 |
9.4 |
3.16 |
|
ƒѕƒµЁп |
Artemisia eriopoda Bge. |
3.98 |
14.2 |
15 |
3.8 |
2.01 |
8 |
103.4 |
3.42 |
|
јдЁп |
Artemisia feddei Levl. |
3.99 |
12.2 |
7.6 |
2.96 |
|
|
|
|
|
ѕЄЅ—“ґЁп |
Artemisia tanacetifolia L. |
25.6 |
1.3 |
0.1 |
0.77 |
3.42 |
6 |
40.4 |
3.78 |
|
“р≥¬Ёп |
Artemisia capillaris Thunb. |
8.1 |
0.7 |
0.6 |
0.57 |
5.25 |
0.33 |
0.2 |
0.23 |
|
√…є≈Ёп |
Artemisia mongolica Fisch. |
4.4 |
1.0 |
0.9 |
0.45 |
2.72 |
2 |
1.9 |
1.26 |
|
÷н√ЂЁп |
Artemisia scoparia Wald. |
|
|
|
|
3.48 |
5 |
25.6 |
2.02 |
|
∆—єЂ”Ґ |
Taraxacum mongolicum |
|
|
|
|
3.18 |
14.7 |
50.4 |
6.05 |
|
∞Ґґыћ©єЈЌёї® |
Heteropappus altaicus Nov. |
9.4 |
1 |
0.5 |
0.91 |
4.90 |
0.33 |
0.2 |
0.30 |
|
њаЁ§≤Ћ |
Ixeris sonchifolia Hance. |
|
|
|
|
3.80 |
0.67 |
0.3 |
0.26 |
|
їр»ё≤Ё |
Leontopodium leontopodioides |
5.7 |
2.3 |
0.3 |
0.54 |
|
|
|
|
|
≤ЁµЎЈз√ЂЊ’ |
Saussurea amara DC. |
|
|
|
|
2.64 |
5 |
14.6 |
1.71 |
|
µє”р“ґЈз√ЂЊ’ |
Saussurea runicinata DC. |
|
|
|
|
3.34 |
5 |
9.1 |
2.39 |
|
ґм»ёќЃЅк≤Ћ |
Potentilla anserina L. |
|
|
|
|
3.23 |
3.33 |
34.4 |
2.71 |
|
ґюЅ—ќЃЅк≤Ћ |
Potentilla bifurca L. |
13.0 |
1.2 |
0.1 |
0.54 |
2.01 |
4.67 |
7.8 |
1.77 |
|
ґаЊ•ќЃЅк≤Ћ |
Potentilla multicaulis Bge. |
17.4 |
2.7 |
0.7 |
2.37 |
3.8 |
1.67 |
0.1 |
0.45 |
|
µЎ”№ |
Sanguisorba officinalis L. |
|
|
|
|
3.40 |
0.33 |
0.1 |
0.17 |
|
…љЊ¬(…љі–) |
Allium senescene L. |
9.6 |
1.7 |
0.7 |
1.03 |
6.78 |
0.67 |
1 |
0.55 |
|
ѕЄ“ґЊ¬ |
Allium tenuissimum L. |
12.5 |
3.7 |
2.9 |
2.25 |
7.57 |
4.67 |
14.6 |
2.70 |
|
±вё£ґє |
Medicago ruthenica L. |
4.8 |
3.3 |
0.7 |
1.0 |
3.00 |
3.33 |
1 |
1.66 |
|
÷±ЅҐї∆№ќ |
Astragalus adsurgens pall. |
25.6 |
1.2 |
0.1 |
0.91 |
3.17 |
3.67 |
0.9 |
2.12 |
|
≈ы’л“ґї∆ї™ |
Thermopsis lanceolata R. Br. |
20.5 |
0.7 |
0.1 |
0.64 |
|
|
|
|
|
ї™±±—“ї∆№ќ |
Hedysarum gmelinii Ledeb. |
25.6 |
1.3 |
0.1 |
1.3 |
|
|
|
|
|
ї∆√ЂЉђґє |
Oxytropis ochrantha Bunge |
13.3 |
1.3 |
0.4 |
0.64 |
|
|
|
|
|
ґј––≤Ћ |
Lepidium apetalum Willd. |
|
|
|
|
3.59 |
3.67 |
4.6 |
1.87 |
|
«Ў№і |
Gentiana macrophylla Pall. |
|
|
|
|
4.18 |
1.33 |
0.9 |
1.03 |
|
ј«ґЊ |
Stellera chamaejasme L. |
|
|
|
|
6.45 |
0.67 |
0.2 |
0.65 |
|
¬нЁю |
Iris ensata Thunb. |
|
|
|
|
11.0 |
5 |
1.9 |
3.96 |
|
¬ыЊ•”ђ„”≤Ё |
Silene repens Patr. |
13.8 |
1.3 |
0.5 |
1.1 |
8.10 |
0.33 |
0.1 |
0.32 |
|
ѕЄ“ґ∆≈∆≈ƒ… |
Veronica linariifolia Pall. |
|
|
|
|
1.59 |
1 |
0.5 |
0.26 |
|
іпќЏјп–Њ∞≈ |
Cymbaria dahurica L. |
4.7 |
1.3 |
1.5 |
0.47 |
|
|
|
|
|
≤сЇъ |
Bupleurum chinensis DC. |
8.0 |
0.3 |
0.1 |
0.31 |
3.62 |
0.67 |
0.4 |
0.39 |
|
ЈјЈз |
Saposhmikovia divaricata S. |
7.47 |
9.2 |
0.3 |
1.2 |
|
|
|
|
|
Љо≈о |
Suaeda glauca Bge. |
|
|
|
|
5.85 |
2 |
1.7 |
1.06 |
|
“∞Їъ¬й |
Linum stelleroides L. |
17.0 |
0.7 |
0.3 |
0.59 |
|
|
|
|
|
Ѕљ∆№ё§ |
Polygonum amphibium L. |
16.5 |
0.3 |
0.1 |
0.49 |
8.5 |
1.33 |
0.4 |
0.87 |
|
Ї„ ≠ |
Lappula myosotis V. |
21.5 |
0.7 |
0.2 |
0.58 |
13.9 |
0.67 |
0.6 |
1.00 |
|
ћ∆Ћ…≤Ё |
Thalictrum petaloideum L. |
14.5 |
0.7 |
0.2 |
1.0 |
|
|
|
|
|
њйЊ•≤ЏЋ’ |
Phlomis umbrosa Turcz. |
13.0 |
2.3 |
0.4 |
1.5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.2 Ј≈ƒЅµЎ”лЄо≤ЁµЎ÷÷„””к±»љѕ
Єо≤ЁµЎ÷÷„””кЅњќ™13968.6Ѕ£/©O£ђЈ≈ƒЅµЎќ™1222.6Ѕ£/©O°£Єо≤ЁµЎ÷÷„””к÷–Їћ±Њњ∆ЇЌЊ’њ∆÷≤ќп÷÷„”’ЉЊшіуґа э£ђЈ÷±рќ™31.7£•ЇЌ33.3£•£їЈ≈ƒЅµЎ÷÷„””кЇћ±Њњ∆ЇЌЊ’њ∆÷≤ќп÷÷„”“≤’ЉЊшіу≤њЈ÷£ђЈ÷±рќ™36.3£•ЇЌ50.6£•°£Єо≤ЁµЎ÷÷„””к√№ґ»„оіуµƒ«∞ќе÷÷÷≤ќпЈ÷±рќ™£ЇƒѕƒµЁп°Ґ≤Џ“ю„”≤Ё°ҐЈјЈз°Ґ≤сЇъЇЌ—т≤Ё°£Ј≈ƒЅµЎ÷÷„””к√№ґ»„оіуµƒ«∞ќе÷÷÷≤ќпЈ÷±рќ™£Ї≤Џ“ю„”≤Ё°Ґ∆—єЂ”Ґ°ҐƒѕƒµЁп°Ґ—т≤ЁЇЌј«ґЊ£ђљб÷÷ЅњґЉ–°”ЏЄо≤ЁµЎ°£
±н1-26 Ј≈ƒЅµЎ°ҐЄо≤ЁµЎ÷÷„””к√№ґ»
|
ќп÷÷ |
…ъїо–Ќ |
÷÷„””к√№ґ»£®Ѕ£/©O£© |
|
Єо≤ЁµЎ |
Ј≈ƒЅµЎ |
|
—т≤Ё |
Leymus chinesis Tzvel. |
PH |
1162.15 |
87.01 |
|
≤Џ“ю„”≤Ё |
Poa annua L. |
PH |
3243.71 |
336.61 |
|
’л√© |
Stipa grandis P. |
PH |
20 |
17.5 |
|
±вЋл±щ≤Ё |
Agropyron cristatum (L.) Gaertn |
PH |
6.2 |
2.1 |
|
ћ¶≤Ё |
Carex cranaocarpa Mey. |
PH |
|
30.7 |
|
ƒѕƒµЁп |
Artemisia eriopoda Bge. |
PH |
3690.7 |
265.25 |
|
јдЁп |
Artemisia feddei Levl. |
SS |
5.4 |
0.28 |
|
ѕЄЅ—“ґЁп |
Artemisia tanacetifolia L. |
PH |
28.2 |
11.76 |
|
“р≥¬Ёп |
Artemisia capillaries Thunb. |
PH |
500.2 |
|
|
√…є≈Ёп |
Artemisia mongolica Fisch. |
PH |
|
33.55 |
|
÷н√ЂЁп |
Artemisia scoparia Wald. |
ABH |
|
19.63 |
|
∆—єЂ”Ґ |
Taraxacum mongolicum |
PH |
|
268.5 |
|
∞Ґґыћ©єЈЌёї® |
Heteropappus altaicus Nov. |
PH |
29.4 |
11.6 |
|
њаЁ§≤Ћ |
Ixeris sonchifolia Hance. |
PH |
|
7.5 |
|
їр»ё≤Ё |
Leontopodium leontopodioides |
PH |
396.9 |
|
|
ґюЅ—ќЃЅк≤Ћ |
Potentilla bifurca L. |
PH |
53.8 |
|
|
ґаЊ•ќЃЅк≤Ћ |
Potentilla multicaulis Bge. |
PH |
45.6 |
|
|
…љЊ¬(…љі–) |
Allium senescene L. |
PH |
261 |
|
|
ѕЄ“ґЊ¬ |
Allium tenuissimum L. |
PH |
122.5 |
|
|
±вё£ґє |
Medicago ruthenica L. |
PH |
111.3 |
4.7 |
|
÷±ЅҐї∆№ќ |
Astragalus adsurgens pall. |
PH |
21.9 |
|
|
≈ы’л“ґї∆ї™ |
Thermopsis lanceolata R. Br. |
PH |
4.5 |
|
|
ї™±±—“ї∆№ќ |
Hedysarum gmelinii Ledeb. |
PH |
85.1 |
|
|
ї∆√ЂЉђґє |
Oxytropis ochrantha Turcz. |
PH |
3.1 |
|
|
ґј––≤Ћ |
Lepidium apetalum Willd. |
AH |
|
14.4 |
|
–°ї®ї®∆мЄЋ |
Dontostemon micranthus C. A. Mey. |
AH |
108.8 |
|
|
«Ў№і |
Gentiana macrophylla Pall. |
PH |
|
|
|
ј«ґЊ |
Stellera chamaejasme L. |
PH |
30.3 |
74.3 |
|
¬нЁю |
Iris ensata Thunb. |
PH |
0.9 |
18.5 |
|
¬ыЊ•”ђ„”≤Ё |
Silene repens Patr. |
ABH |
4.1 |
|
|
іпќЏјп–Њ∞≈ |
Cymbaria dahurica L. |
PH |
19.4 |
|
|
≤сЇъ |
Bupleurum chinensis DC. |
PH |
1814.4 |
|
|
ЈјЈз |
Saposhmikovia divaricata S. |
PH |
2113.4 |
|
|
Љо≈о |
Suaeda glauca Bge. |
AH |
|
1.5 |
|
“∞Їъ¬й |
Linum stelleroides L. |
ABH |
|
|
|
Ѕљ∆№ё§ |
Polygonum amphibium L. |
PH |
4.9 |
|
|
Ї„ ≠ |
Lappula myosotis V. |
AH |
35.7 |
17.2 |
|
∞к»пћ∆Ћ…≤Ё |
Thalictrum petaloideum L. |
PH |
7.9 |
|
|
њйЊ•≤ЏЋ’ |
Phlomis umbrosa Turcz. |
PH |
37.2 |
|
|
„№Љ∆£Ї |
13968.66 |
1222.59 |
|
|
|
|
|
|
„Ґ: AH: “їƒк…ъ≤Ё±Њ; ABH: “їґюƒк…ъ≤Ё±Њ: BH: ґюƒк…ъ≤Ё±Њ; PH: ґаƒк…ъ≤Ё±Њ; SS: ∞лєаƒЊ(–°∞лєаƒЊ)
5.3 Ј≈ƒЅµЎ”лЄо≤ЁµЎ÷÷„”њв±»љѕ
Єо≤ЁµЎ°ҐЈ≈ƒЅµЎЌЅ»ј÷÷„”њвіҐЅњµƒ ±ЉдґѓћђЈ÷±рќ™747.7°Ђ1505.4Ѕ£/©OЇЌ303.0°Ђ520.7Ѕ£/©O£їЄо≤ЁµЎ÷÷„”њвЈ÷≤гґѓћђ£®њ’Љдґѓћђ£©Ј÷±рќ™£Ї0°Ђ2©MЌЅ≤г÷÷„”√№ґ»445.0°Ђ833.2Ѕ£/©O°Ґ2°Ђ5©MЌЅ≤г÷÷„”√№ґ»170.4°Ђ463.9Ѕ£/©OЇЌ5°Ђ10©MЌЅ≤г÷÷„”√№ґ»132.6°Ђ208.3Ѕ£/©O£ђЈ≈ƒЅµЎ÷÷„”њвЈ÷≤гґѓћђЈ÷±рќ™£Ї0°Ђ2©MЌЅ≤г÷÷„”√№ґ»179.9°Ђ340.8Ѕ£/©O°Ґ2°Ђ5©MЌЅ≤г÷÷„”√№ґ»85.2°Ђ161.0Ѕ£/©OЇЌ5°Ђ10©MЌЅ≤г÷÷„”√№ґ»18.9°Ђ28.4Ѕ£/©O°£Єо≤ЁµЎ°ҐЈ≈ƒЅµЎ÷÷„”њв÷ч“™Љѓ÷–‘Џ0°Ђ5©MЌЅ≤г÷–£ђ”…«≥»л…оґшµЁЉх°£Ј≈ƒЅњ… єЊ’њ∆÷≤ќп÷÷„”±»јэ‘ціу°£÷÷„”њв÷–Њ’њ∆÷≤ќп÷÷„”ЈҐ—њЋўґ»“™њм”ЏЇћ±Њњ∆ЇЌґєњ∆£ђЋщ“‘‘≠”–÷≤±ї∆∆їµ“‘Їу£ђЊ’њ∆÷≤ќп ∆±Ў“™’ЉЅмњ’Љд£ђµЉ÷¬ƒЅ≤Ё÷ Ѕњѕ¬љµ°£
ЅнЌвЄо≤Ёґ‘≤ЁµЎ÷≤ќпµƒЄ…»≈≥ћґ»–°”ЏЈ≈ƒЅґ‘≤ЁµЎ÷≤ќпµƒЄ…»≈£їЈ≈ƒЅµЉ÷¬÷≤÷к–°–Ќїѓ°ҐЄщЊ•љЏЉдЊајлЋхґћ£їЌђ ±£®«бґ»£©Ј≈ƒЅ“≤њ…“‘ћбЄя≤ЁµЎ÷≤ќпґа—щ–‘°£
6°Ґ≤ЁµЎ≤ЁµЎ÷≤±їґ‘љµ”кЅњ±дїѓµƒѕм”¶
±Њ ‘—й”Џ2005ƒк6‘¬‘ЏЇ”±±∞”…ѕ’≈Љ“њЏєЅ‘іƒЅ≥°—т≤Ё≤Ё‘≠£ђљш––ЅЋљµ”кЅњµƒ≤®ґѓЇЌјы”√«њґ»±дїѓµƒ ‘—йі¶јн°£љµ”кЅњ≤®ґѓµƒі¶јн «≤…”√’Џ”к…и±ЄјєљЎ50£• ‘—й–°«ш0.5m’э…ѕњ’µƒ√жїэ£ђ“‘іпµљЉх…ў50£•љµЋЃЅњµƒ“™«у£ђ ‘—й µЉ ≤вґ®µƒјєљЎ–Ієы‘Џ43£•£≠48£•÷ЃЉд£ђ‘цЉ”50£•і¶јнµƒ ‘—й–°«ш≤…”√»Ћє§љљЋЃµƒ∞мЈ®іпµљ“™«у°£ЄщЊЁ∆шѕу’Њµƒ≤вґ®љбєы£ђљш––љљЋЃ°£јы”√«њґ»…и÷√ЅљЄцћЁґ»£ђ“їЄц»ЂƒкЎ„Єо2іќ£ђ“їЄцљцЎ„Єо“їіќ£©°£ґ‘ ‘—йі¶јн÷–µƒ—т≤ЁµЎ…ѕµЎѕ¬…ъќпЅњ°Ґ ‘—йі¶јн≤їЌђ≤гіќЌЅ»јЇђЋЃЅњƒкґ»±дїѓ°Ґ—т≤Ё»Ї¬д„й≥…“‘Љ∞≤їЌђ„йЈ÷≤ъЅњµƒЈ÷≤Љµ»љш––ЅЋ—–Њњ°£
6.1 љµЋЃЅњ±дїѓЇЌјы”√«њґ»ґ‘µЎ…ѕ…ъќпЅњ±дїѓµƒ”∞ѕм
Ћжјы”√«њґ»µƒ‘цЉ”£ђ≤ЁµЎµƒµЎ…ѕ…ъќпЅњ”–љµµЌµƒ«ч ∆£ђ‘цЉ”љµЋЃЅњЇЌЉх…ўљµ”кЅњґЉ“їґ®≥ћґ»µƒЉх…ўЅЋ—т≤Ё»Ї¬дµƒ…ъќпЅњµƒ«ч ∆£ђЈљ≤оЈ÷ќц≤їѕ‘÷ш°£Ў„Єо«њґ»‘цЉ”Ќђ ±љµ”кЅњЉх…ўµƒ«йњцѕ¬£ђ…ъќпЅњѕ‘÷шљµµЌ£ї‘Џ’э≥£ЇЌ‘цЉ”50£•љµЋЃЅњі¶јн÷–‘цЉ”јы”√«њґ»£ђ…ъќпЅњљµµЌ≤Ґ≤ї√чѕ‘°£Ћµ√чјы”√«њґ»µƒ‘цЉ”їбЉх…ўµЎ…ѕ…ъќпЅњ£ђ”»∆д «Є…ЇµµƒƒкЈЁїбљш“ї≤љЉ”ЊзµЎ…ѕ…ъќпЅњµƒЉх…ў£®ЌЉ1-10£©°£
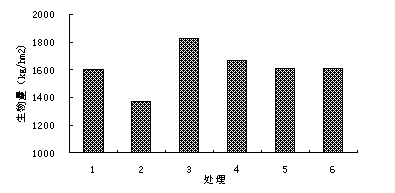
ЌЉ1-10 ≤їЌђі¶јнЉдƒк…ъќпЅњѕм”¶£®2005ƒк£©
„Ґ£Їі¶јн1±н Њљµ”кЅњЉх…ў50£•£ђ»ЂƒкЎ„Єо1іќ£їі¶јн2±н Њљµ”кЅњЉх…ў50£•£ђ»ЂƒкЎ„Єо2іќ£їі¶јн3±н Њљµ”кЅњ≤ї±д£ђ»ЂƒкЎ„Єо1іќ£їі¶јн4±н Њљµ”кЅњ≤ї±д£ђ»ЂƒкЎ„Єо2іќ£їі¶јн5±н Њљµ”кЅњ‘цЉ”50£•£ђ»ЂƒкЎ„Єо1іќ£їі¶јн6±н Њљµ”кЅњ‘цЉ”50£•£ђ»ЂƒкЎ„Єо2іќ°£
6.2 ƒкЉ Љдљµ”кЅњ±дїѓґ‘—т≤Ё»Ї¬дµЎ…ѕ…ъќпЅњ“‘Љ∞»Ї¬д„й≥…µƒ”∞ѕм
”…ЌЉ1-11њ…“‘њі≥ц2006ƒкµƒ…ъ≥§ЉЊµƒљµЋЃЅњќ™311.79mm±»2005ƒкµƒ216.7‘цЉ”ЅЋ95.09mm.—т≤Ё»Ї¬дµƒ…ъќпЅњ√чѕ‘‘цЉ””…2005ƒкµƒ159g/m2 ‘цЉ”µљ 191g/m2іпµљѕ‘÷ш–‘≤о“м£ђ—т≤Ёµƒ≤ъЅњ”…112g/m2Љх…ўµљ100g/m2іпµљѕ‘÷ш–‘≤о“м°£
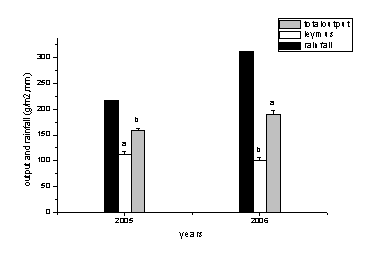
ЌЉ1-11 љµ”кЅњЇЌµЎ…ѕ…ъќпЅњƒкґ»Љд±дїѓ
”…2005ЇЌ2006ƒк≤їЌђі¶јнЉд—т≤Ё»Ї¬дљ®»Ї÷÷„й≥…µƒ±дїѓњ…“‘њі≥ц£ђЋж„≈љµ”кЅњµƒ‘цЉ”£ђ‘Џ≤ъЅњєє≥……ѕƒѕƒµЁпЇЌ≤Џ“ю„”≤Ёµƒ±»јэ‘цЉ”£ђґш—т≤Ёµƒ≤ъЅњ≥ ѕ÷ѕ¬љµµƒ«ч ∆°£Ј÷ќц÷ч“™µƒ‘≠“т «ƒѕƒµЁпЇЌ≤Џ“ю„”≤ЁµƒЄщѕµ÷ч“™Ј÷≤Љ‘ЏЌЅ»ј±н≤г0£≠8cm’вЄц≤гіќ£ђ—т≤ЁµƒЄщЊ•Љѓ÷–‘Џ8£≠14cm°£љµЋЃЅњµƒ‘цЉ”ћбЄяЅЋЌЅ»ј±н≤гµƒЇђЋЃЅњ£ђ‘цЉ”ЅЋƒѕƒµЁпЇЌ≤Џ“ю„”≤Ё‘Џ÷÷»Ї÷–µƒЊЇ’щƒ№Ѕ¶£ђ“тіЋ—т≤Ёµƒ…ъќпЅњ‘ЏљµЋЃЅњ‘цЉ”µƒ«йњцѕ¬Јіґш≥цѕ÷ЅЋѕ¬љµµƒѕ÷ѕу£®ЌЉ1-12£ђЌЉ1-13£©°£

ЌЉ1-12 2005ƒк—т≤Ё»Ї¬дљ®»Ї÷÷µƒ≤ъЅњєє≥…

ЌЉ1-13 2006ƒк—т≤Ё»Ї¬дљ®»Ї÷÷µƒ≤ъЅњєє≥…
6.3јы”√«њґ»ґ‘—т≤Ё÷÷»Їµƒ”∞ѕм
јы”√«њґ»ґ‘—т≤Ё»Ї¬дљ®»Ї÷÷µƒ”∞ѕмЇЌљµ”кЅњµƒіу–°”–√№«–µƒєЎѕµ£ђ”…ЌЉ1-14њ…“‘њі≥ц£ђ2006ƒкљµЋЃЅњ‘цЉ”£ђјы”√«њґ»‘цЉ”Їу—т≤Ёµƒ…ъќпЅњ≥ ѕ÷‘цЉ”µƒ«ч ∆£їґш2005ƒкљµ”кЅњљѕµЌ£ђЋжјы”√«њґ»‘цЉ”Їу£ђ—т≤Ё≤ъЅњ≥цѕ÷ѕ¬љµµƒ«ч ∆£ђ‘Џ2005°Ґ2006ƒкљµ”кЅњ±дїѓ єƒѕƒµЁпЇЌ≤Џ“ю„”≤Ёµƒ„№µЎ…ѕ…ъќпЅњ”…33£≠40g/m2‘цЉ”µљ72g/m2≤ъЅњ√чѕ‘‘цЉ”£їґшјы”√«њґ»і¶јнґ‘2005ЇЌ2006ƒк—т≤Ё»Ї¬д÷–µƒƒѕƒµЁпЇЌ≤Џ“ю„”≤Ёµƒ”∞ѕм≤їіу£®ЌЉ1-14£©°£

ЌЉ1-14 Ў„Єо«њґ»ґ‘—т≤Ё÷÷»Ї„й≥…µƒЇЌµЎ…ѕ…ъќпЅњµƒ”∞ѕм
6.4 љµ”кЅњ±дїѓґ‘≤ЁµЎ…ъ≥§Ћў¬ µƒ”∞ѕм
2006ƒк—т≤Ё»Ї¬дµƒ„№≤ъЅњ±»2005ƒкћбЄяЅЋ48.9£•£ђ—т≤Ё≤ъЅњћбЄя43.9£•£ђ2006ƒк…ъ≥§ЉЊЄчЄц ±∆ЏµƒµЎ…ѕ…ъќпЅњґЉЄя”Џ2005ƒкµƒѕа”¶µЎ…ѕ…ъќпЅњ°£2006ƒк…ъ≥§ЉЊµƒљµ”кЅњќ™318mm£ђ‘Џ8‘¬20»’«∞љµ”к±»љѕЊщ‘»£ђі”ЌЉ÷–њ…“‘њі≥ц2006ƒк—т≤Ё»Ї¬д„№…ъќпЅњЇЌ—т≤Ёµƒ…ъќпЅњґЉ «÷рљ•‘цЉ”µƒ£ђґш2005ƒк…ъ≥§ЉЊµƒљµ”кќ™200mmґш«“…ъ≥§ЉЊ«∞∆Џ∆ЂЇµ£ђ…ъќпЅњµƒїэјџіж‘Џ√чѕ‘µƒЌ£÷є£ђЈіµѓµƒѕ÷ѕу£®ЌЉ1-15£©°£
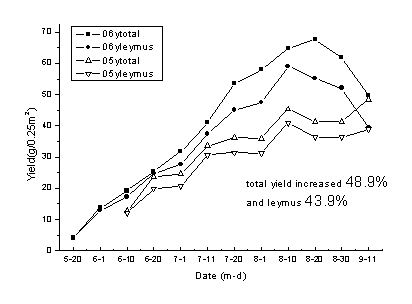
ЌЉ 1-15 2005ƒкЇЌ2006ƒк≤їЌђ ±∆Џ—т≤Ё”л»Ї¬д„№≤ъЅњµƒµЎ…ѕ…ъќпЅњ±дїѓ
ґ‘2006ƒк…ъ≥§Ћў¬ £®ЌЉ1-16£©љш––Ј÷ќц±н√ч£ђ8‘¬20»’«∞’ыЄц»Ї¬д «’э‘ц≥§£ђ8‘¬20»’„у”“»Ї¬дµƒµЎ…ѕ…ъќпЅњіпµљЈе÷µ£ђљ”„≈»Ї¬дµƒµЎ…ѕ…ъќпЅњњ™ Љѕ¬љµ°£÷µµ√„Ґ“вµƒ «8‘¬1»’’в“їµг£ђі”ЌЉ÷–њ…“‘њі≥ц’в“їґќ ±Љд…ъ≥§Ћў¬ √чѕ‘љµµЌ£ђі” ‘—йµЎ÷–—лµƒ ‘—й’Њµ√µљµƒ∆шѕу эЊЁ «љµ”кЅњ34.4mm£ђ∆љЊщќ¬ґ»17.7°ж£ђћЂ—фЈш…д183.3w/m2£ђЈзЋў6.4m/s£ђЌЅ»јЇђЋЃЅњ13.0£•£ї7‘¬20»’µƒ эЊЁЈ÷±рќ™66.3°Ґ19.4°Ґ242.3°Ґ73.9°Ґ7.3°Ґ14.3£ї8‘¬10»’µƒќ™27.5°Ґ20.0°Ґ233.2°Ґ84.4°Ґ5.0°Ґ11.0°£њ…“‘њі≥ц÷ч“™µƒ«ш±р‘Џ”л∆љЊщќ¬ґ»°ҐћЂ—фЈш…дЇЌѕаґ‘ ™ґ»£ђ”…”Џ’вґќ ±ЉдћЂ—фЈш…дµЌ£ђ’фЈҐЅњ–°њ’∆шµƒѕаґ‘ ™ґ»љѕЄя£ђњ’∆шќ¬ґ»љѕµЌ£ђ“тіЋ…ъ≥§Ћў¬ ЉхїЇ°£”…іЋњ…ЉыЉі є‘Џ…ъ≥§Ќъ ҐµƒЉЊљЏ£ђ‘Џ∞лЄ…ЇµµЎ«шљµ”кЅњ“≤≤ї «ќ®“їЊцґ®“тЋЎ£ђ’в“їЈҐѕ÷Љдљ”µƒїЎірЅЋќ™ ≤√і‘Џ”√љµ”кЅњ”л…ъќпЅњљш––ѕаєЎЈ÷ќц ±£ђ≤о“м≤Ґ≤ї“їґ®ѕ‘÷шµƒќ ћв°£

ЌЉ1-16 ≤їЌђ ±∆Џ—т≤ЁЇЌ»Ї¬дµƒ…ъ≥§Ћў¬
6.5 љµЋЃЅњ±дїѓЇЌјы”√«њґ»ґ‘µЎѕ¬…ъќпЅњµƒ”∞ѕм
‘—йі¶јн÷–Ћж„≈љµ”кЅњµƒ‘цЉ”µЎѕ¬…ъќпЅњ”–Љх…ўµƒ«ч ∆£®ЌЉ1-17£©£ђЋж„≈Ў„

ЌЉ1-17 љµ”кЅњЇЌјы”√«њґ»ґ‘µЎѕ¬…ъќпЅњµƒ”∞ѕм
„Ґ£ЇBNPP±н ЊµЎѕ¬…ъќпЅњ£ђrhizome±н Њ—т≤ЁµЎѕ¬ЄщЊ•£ђroots±н Њ∆дЋьЄщѕµ°£
Єоіќ эµƒ‘цЉ”‘ЏЉх…ў50£•µƒі¶јнЇЌљµ”кЅњ≤ї±дµƒі¶јн÷–µЎѕ¬…ъќпЅњ”–‘цЉ”µƒ«ч ∆°£‘Џ2005ƒк…ў”кƒкЇЌ2006ƒкЈб”кƒкµƒ±»љѕ÷–£ђ2006ƒкµƒ÷≤ќпЄщѕµ„№Ѕњ“™Єя”Џ2005ƒкµƒЋЃ∆љ£ђ’в“їљбєыЇЌƒкƒЏµƒљµ”кЅњ±дїѓ’эЇ√ѕаЈі£ђ‘ЏЈљ≤оЈ÷ќц÷–≤о“мќііпµљѕ‘÷шЋЃ∆љ°£
6.6 –°љб
‘Џ∞лЄ…Їµ«ш≤Ё‘≠£ђ„чќ™÷ч“™„‘»їѕё÷∆“т„”µƒљµЋЃЅњґ‘—т≤Ё»Ї¬дµƒµЎ…ѕ…ъќпЅњ”∞ѕмЇ№іу£ђ±Њ ‘—й÷–2006ƒк„№≤ъЅњ±»2005ƒк‘цЉ”ЅЋ48.9£•£ђ“тіЋќ“√«≤їƒ№“‘Єц±рƒкЈЁ≤Ё‘≠…ъќпЅњљµµЌ„чќ™≈–ґ®≤Ё≥°ЌЋїѓµƒ÷Є±к°£“тќ™‘Џ„‘»їћхЉюѕ¬£ђƒкЉ Љд №љµ”кЅњµƒ”∞ѕм£ђ≤Ё≥°…ъќпЅњіж‘ЏЇ№іуµƒ≤®ґѓ–‘°£
‘—йљбєы“≤Ћµ√чќ“√«‘Џґ‘≤Ё‘≠љш––є№јн ±£ђ»з‘Џљш––≤Ё–у∆љЇв£ђ“‘≤Ёґ®–у ±–и“™њЉ¬«„‘»їћхЉюµƒ±дїѓ£ђ”»∆д «‘Џ∞лЄ…Їµ≤Ё‘≠«ш°£
‘Џ∞лЄ…ЇµµЎ«ш£ђ÷≤ќпЌъ Ґ…ъ≥§µƒ ±∆Џ»зєыћЂ—фЈш…дЇЌќ¬ґ»љµµЌљЂљµµЌƒЅ≤Ёµƒ…ъ≥§Ћў¬ °Ґѕё÷∆≤ЁµЎµƒ…ъќпЅњµƒїэјџ°£
7£ЃЌЋїѓ≤ЁµЎї÷ЄіЄƒЅЉƒ£ љ
Є Ћа≤Ё‘≠…ъћђ—–ЊњЋщ“‘≈©“µ≤њ≤ЁµЎ≈©“µ…ъћђ—І÷Ўµгњ™Ј≈ µ—й “£®23260271£©ЇЌ«м—фї∆ЌЅЄя‘≠≤ЁµЎ≈©“µѕµЌ≥ ‘—й’Њќ™“јЌ–£ђ”Џ2002---2006ƒк‘ЏЄ ЋаїЈѕЎљш––ЅЋЌЋїѓ≤ЁµЎ„џЇѕЄƒЅЉЉЉ х—–Њњ£ђ’лґ‘≤їЌђЌЋїѓљ„ґќћм»ї≤ЁµЎћб≥цЅЋ≤їЌђµƒЄƒЅЉЉЉ х°£
ћб≥цЅЋ “Ћ”Џ÷Ўґ»ЌЋїѓ≤ЁµЎµƒ°∞Јв”э+їЃ∆∆+≤є≤•°±„џЇѕЄƒЅЉ°£ґ‘±» ‘—й±н√ч£ђЈв”э°Ґ≤є≤•°ҐїЃ∆∆≤Ё∆§Њщƒ№єї єЌЋїѓ≤ЁµЎµ√“‘ї÷Єі£ђµЂ3÷÷ЈљЈ®„џЇѕЉѓ≥…Їу£ђЊя”–Љтµ•°Ґ“„≤ў„ч°Ґ–Ієы√чѕ‘µ»ћЎµг£ђ«– µњ…––£ђ ”Џ ‘—й«шЌ∆єг°£
ћб≥цЅЋ ”Џ÷–ґ»ЌЋїѓ≤ЁµЎµƒ°∞ЅљЉЊїЃ«ш¬÷ƒЅ°±ЄƒЅЉЉЉ х°£∞—≤ЁµЎЈ÷ќ™≈ѓЉЊЈ≈ƒЅ«шЇЌјдЉЊЈ≈ƒЅ«шЅљја£ђЈ÷±р‘Џјд°Ґ≈ѓЅљЉЊїЃ«ш¬÷ƒЅ£ђ„џЇѕЌЅ°Ґ≤Ё°Ґ–ућЎ’ч£ђ»Јґ® “Ћµƒ‘Ў–уЅњ£ђ≤Ґ”√°∞г–ѕёЋЂ“т„”Ј®°±÷ЄµЉЄч¬÷ƒЅ–°«шµƒЈ≈ƒЅњ™ ЉЇЌљб ш ±Љд°£Є√ƒ£ љ£ђЉ»Ќ®єэЉ“–уЈ≈ƒЅ±£≥÷’э≥£µƒ≤Ё–уєЎѕµ£ђќђї§≤ЁµЎљ°њµ£ђ”÷њ…“‘”лґћ∆ЏЈ ”эљбЇѕ…ъ≤ъЅљЉЊ–у≤ъ∆Ј£ђЄƒ…∆“‘ЌщЉ“–уЈ ”э÷ч“™Љѓ÷–”ЏјдЉЊµƒµ•“їƒ£ љ°£
ћб≥цЅЋ°∞ЌЋЄыµЎЈв”э°±µƒ„‘»її÷ЄіЉЉ х°£ЌЋЄыµЎЊ≠єэЄы„ч£ђЋЃЈ÷ћхЉюЇ√£ђЌЅ»јЈ Ѕ¶Єя£ђ≤ъ≤ЁЅњ‘Џї÷Єі≥х∆ЏЇ№Єя£ђќь“эЉ“–уЈ≈ƒЅ£ђµЂ≥х∆Џ÷ч“™…ъ≥§“їƒк…ъƒЅ≤Ё£ђЈ≈ƒЅµЉ÷¬ƒЅ≤Ё…ъ≥§≤ї≥дЈ÷£ђƒ—“‘–ќ≥…Ј±÷≥∆чєў£ђЌЅµЎ—ЄЋўЌЋїѓ£ђ≤…”√Јв”эЇу£ђ ЃЉЄƒкЊЌњ…“‘—ЄЋўї÷Єіµљљ°њµЋЃ∆љ°£
‘—й«шћм»ї≤ЁµЎ÷≤±їЄ«ґ»ї÷Єіµљ90%“‘…ѕ£ђµЎ…ѕ…ъќпЅњћбЄя1±ґ„у”“°£‘Џ…ѕ цЉЉ хµƒ÷І≥≈ѕ¬£ђїЈѕЎћм»ї≤Ё‘≠ЄƒЅЉ ЊЈґ”лЌ∆єг√жїэ≥ђєэ2600hm2°£Ќ®єэ≤є≤•£ђїЃ∆∆µ»Јљ љї÷Єіћм»ї≤Ё‘≠500 hm2“‘…ѕ°£ї÷Єі«ш÷≤±їЄ≤Є«¬ іпµљ75%“‘…ѕ£ђ ЊЈґ«шЌЅ»ј”–їъ÷ ЇђЅњ∆љЊщћбЄя0.2%“‘…ѕ°£‘Ў–уЅњ3600—тµ•ќї£ђ÷№±яѕз’т300ґа≈©їІ÷±љ” №“ж°£
7.1 °∞Јв”э+їЃ∆∆+≤є≤•°±„џЇѕЄƒЅЉЉЉ х
‘Џ÷Ўґ»ЌЋїѓµƒ„‘”…Ј≈ƒЅ≤ЁµЎ…ѕљ®ЅҐќІЈв°ҐїЃ∆∆+ќІЈв°ҐќІЈв+≤є≤•°ҐќІЈв+≤є≤•+їЃ∆∆ ‘—й—щµЎ°£≤вґ® ‘—й«∞ЇуЄчі¶јнµƒµЎ…ѕѕ÷іжЅњ°Ґµт¬дќпЇЌќп÷÷ эµ»÷≤±їћЎ’ч°£±»љѕЄч÷÷ЄƒЅЉіл ©µƒ”≈µгЇЌ»±µг£ђ≤Ґґ‘єЎЉьЉЉ хњ™’є„®ѕо—–Њњ°£
7.1.1 ґ‘≤ЁµЎµЎ…ѕѕ÷іжЅњ°Ґµт¬дќп”лЌЅ»ј”–їъћЉµƒ”∞ѕм
„№ће…ѕ£ђЈв”эњ™ ЉЇу1ƒкƒЏ£ђ÷Ўґ»ЌЋїѓ≤ЁµЎµЎ…ѕѕ÷іжЅњ°Ґµт¬дќп”лЅҐњЁќпЅњЉ±ЊзЉх…ў£ђ÷ч“™“тќ™“їƒк…ъ÷≤ќпЉх…ўљѕґа£ђµЂЌЅ»ј”–їъћЉ‘Џ5ƒкƒЏ÷рљ•‘цЉ”£®ЌЉ1-18£©°£»їґш£ђƒ≥–©µЎ–ќћхЉюѕ¬£ђ»з—ф∆¬15°гЇЌ45°г£ђЈв”эƒ№Јсѕ‘÷ш‘цЉ”≤ЁµЎµЎ…ѕ…ъќпЅњ£ђїє»°Њц”ЏљµЋЃћхЉю°£ѕ÷іжЅњµƒ‘цЈщ£ђ“х∆¬„оЄяќ™25%£ђ∆¬ґ•ќ™20%°™60%£ђ—ф∆¬≤ї≥ђєэ35%£ђ»эЄц∆¬ѕтµƒ±дїѓ«ч ∆ќ™∆¬ґ•>—ф∆¬>“х∆¬°£“х∆¬15°гЇЌ—ф∆¬45°гћхЉюѕ¬£ђЈв”эґ‘≤ъ≤ЁЅњ√ї”–ѕ‘÷ш”∞ѕм£®p>0.05£©°£Јв”эЇу£ђ35%-50%µƒµЎ…ѕќп÷ „чќ™µт¬дќпіж‘Џ”Џ≤ЁµЎ÷–°£≤ЁµЎµт¬дќпЅњґ‘”Џ±£ї§ЌЅ»ј£ђЉхїЇЋЃЌЅЅч І”–÷Ў“™„ч”√£ђЈв”эЇу≤ЁµЎµт¬дќпѕ÷іжЅњ±»ќіЈв”э≤ЁµЎѕ‘÷ш‘цЉ”£®p<0.01£©°£”лЈ≈ƒЅ∆¬µЎѕа±»£ђ‘Џ“х∆¬£ђ15°г°Ґ30°гЇЌ45°гµƒЈв”э∆¬µЎµт¬дќпЅњ‘цЉ”2-4±ґ£ђ∆¬ґ•‘тћбЄя3-15±ґ£ђ—ф∆¬Єя4-15±ґ°£‘Џљѕќ™Є…‘пµƒ—ф∆¬ЇЌ∆¬ґ•£ђЈв”эґ‘”Џµт¬дќпїэјџ”–Єьќ™їэЉЂµƒ„ч”√°£
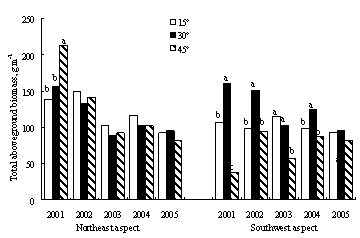

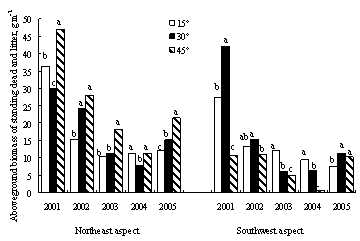
ЌЉ1-18Јв”э≤ЁµЎѕ÷іжЅњ°Ґµт¬дќпЅњ”лЌЅ»ј”–їъћЉЇђЅњ
7.1.2 ґ‘≤ЁµЎ…ъќпЅњ°ҐЄ«ґ»ЇЌќп÷÷ЈбЄїґ»µƒ”∞ѕм
Єч÷÷ЄƒЅЉіл ©ѕ¬£ђƒЅ≤Ё…ъ≤ъЅ¶іу–°“јіќќ™£ЇїЃ∆∆+≤є≤•+Јв”э>≤є≤•+Јв”э>Јв”э>„‘”…Ј≈ƒЅ£®ЌЉ1-19£©°£Јв”э+≤є≤•+їЃ∆∆і¶јнµƒ≤ЁµЎ£ђƒЅ≤ЁµЎ…ѕѕ÷іжЅњ£®225g/m2£©”лµт¬дќпЅњ£®54.93g/m2£©„оЄя£ђЈ÷±р «Јв”э≤ЁµЎµƒ1.9±ґЇЌ10±ґ°£‘ЏЈ≈ƒЅћхЉюѕ¬£ђ”л÷±љ”≤є≤•µƒЄƒЅЉЈљЈ®ѕа±»£ђїЃ∆∆+≤є≤•і¶јнµƒ≤ЁµЎƒЅ≤Ёѕ÷іжЅњ”лµт¬дќпЅњЈ÷±р‘цЉ”0.43±ґ°Ґ6.66±ґ°£Ћµ√ч£ђЌЋїѓ≤ЁµЎµƒЄƒЅЉ”лЈ≈ƒЅ≤Ґ≤їґ‘ЅҐ°£ґћ∆ЏƒЏ£ђ≤їЌђї÷ЄіЈљ љґ‘≤ЁµЎќп÷÷ э°ҐЄ«ґ»”∞ѕм”–Ћщ≤о“м£ђЄя”Џ„‘”…Ј≈ƒЅµЎ20%-30%£®ЌЉ1-19£©°£Ћµ√чЈв”эƒ№єїћбЄя≤ЁµЎµƒ…ъћђ–І“ж°£µЂ «£ђЈв”э≤Ґ≤їѕ‘÷ш‘цЉ”ќп÷÷ЈбЄїґ»£®p>0.05£©°£

ЌЉ1-19 2004ƒкЄч÷÷ї÷Єііл ©ґ‘≤ЁµЎ…ъќпЅњ°ҐЄ«ґ»ЇЌќп÷÷ЈбЄїґ»µƒ”∞ѕм
 

ЌЉ1-20 їЃ∆∆«њґ»ґ‘≤ЁµЎµƒ”∞ѕм
їЃ∆∆«њґ»£®їЃ∆∆«ш”т’Љ’ыЄц≤ЁµЎµƒ√жїэ±»јэ£©ќ™46.3% ±µЎ…ѕ…ъќпЅњ„оЄя£®ЌЉ1-20£©°£µЂ «ЄщЅњ‘Џ±ЊѕоƒњµƒїЃ∆∆«њґ»ЈґќІƒЏ£ђ”л÷Ѓ≥…’э±»£їЌЅ»јЇђЋЃЅњЋж÷Ѓѕ¬љµ£ђµЂїЃ∆∆«њґ»46.3%”л61.9%÷ЃЉдќёѕ‘÷ш≤о“м£®p>0.05£©°£„џЇѕ3ѕо÷Є±к£ђ“‘їЃ∆∆«њґ»≤ї≥ђєэ50%ќ™“Ћ£ђ»зіЋ£ђ≤ЁµЎЉ»Њя”–љѕЄяµƒµЎ…ѕ…ъќпЅњ£ђ“≤Њя”–“їґ®µƒµЎѕ¬…ъќпЅњ°£
≤їЌђќІЈв ±ѕёґ‘≤ЁµЎ…ъ≤ъЅ¶ЇЌќп÷÷ЈбЄїґ»”–ѕ‘÷ш”∞ѕм£®ЌЉ1-21£©°£2005ƒк£ђќІЈв6ƒкµƒ≤ЁµЎ≤ъ≤ЁЅњ±»„‘”…Ј≈ƒЅµЎЄя78.7%£®p<0.01£©£ђ2006ƒкќІЈв3ƒкЇЌ6ƒкµƒ—щµЎ≤ъ
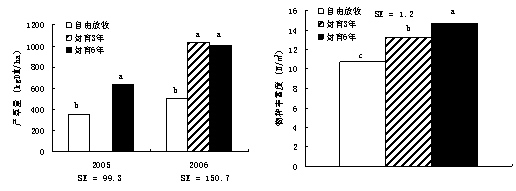
ЌЉ1-21 Ј≈ƒЅЇЌЈв”эґ‘≤ъ≤ЁЅњЇЌќп÷÷ЈбЄїґ»µƒ”∞ѕм
≤ЁЅњ≥ђ≥ц„‘”…Ј≈ƒЅµЎ1±ґ£®p<0.01£©°£2006ƒк£ђќп÷÷ЈбЄїґ»“‘Јв”э6ƒкµƒ—щµЎ„оЄя£ђґш“‘„‘”…Ј≈ƒЅµЎ„оµЌ°£Ћµ√ч£ђ≤ЁµЎ…ъ≤ъЅ¶µƒї÷Єі“™њм”Џќп÷÷„й≥…°£
7.2 °∞ЅљЉЊїЃ«ш¬÷ƒЅ°±ЄƒЅЉЉЉ х
—°‘сµЎ ∆∆љћє°Ґ÷≤±ї“ї÷¬µƒ÷–ґ»ЌЋїѓћм»ї≤ЁµЎ£ђљ®ЅҐћ≤—тїЃ«ш¬÷ƒЅ ‘—й–°«ш20Єц°£ѕƒЉЊ¬÷ƒЅ≤ЁµЎ£ђ√њƒк6‘¬÷–—Ѓ÷Ѕ9‘¬÷–—Ѓ¬÷ƒЅ°£√њ–°«шЅђ–шЈ≈ƒЅ10d£ђ¬÷ƒЅ÷№∆Џ30d£ђ“їƒк¬÷ƒЅ3іќ°£ ‘—йЉ“–уќ™1-2Ћкµƒ»• ∆єЂћ≤—т£ђћењцѕаљь£ђЈ≈ƒЅ«њґ»Ј÷±рќ™0£®ґ‘’’£©°Ґ2.67—т/hm2°Ґ5.33—т/ hm2ЇЌ8.67—т/hm2°£√њ–°«шЈ≈ƒЅ«∞£ђЉ“–уњ’Єє≥∆÷Ў£ђ10ћм≥∆“їіќ°£Ј≈ƒЅ«∞Їу≤вґ®Єч–°«шƒЅ≤Ёѕ÷іжЅњ°Ґµт¬дќпЅњ°Ґќп÷÷ эЇЌ÷≤±їЄ«ґ»£ђ«пЉЊ≤вґ®0-5cmЌЅ≤гЅ£ЊґЇЌ0-30cm”–їъ÷ °£ґђЉЊ¬÷ƒЅ≤ЁµЎ£ђ√њƒк11‘¬÷Ѕ12‘¬÷–—Ѓ¬÷ƒЅ£ђ√њ–°«шЅђ–шЈ≈ƒЅ5d£ђ¬÷ƒЅ÷№∆Џ15d£ђ“їƒк¬÷ƒЅ3іќ°£
7.2.1 ≤ЁµЎЇѕјнЈ≈ƒЅЉЉ х°™»Јґ® “ЋЈ≈ƒЅ«њґ»
‘Џ¬÷ƒЅѕµЌ≥÷–£ђ»Јґ® “ЋµƒЈ≈ƒЅ«њґ» « µ––≤ЁµЎљ°њµє№јнµƒїщі°°£ѕƒЉЊ¬÷ƒЅ£ђ‘Ў–уЅњќ™2.67—т/ha ±£ђ≤ЁµЎЊя”–„оіуµƒµЎ…ѕѕ÷іжЅњ°Ґќп÷÷ЈбЄїґ»ЇЌЌЅ»ј”–їъћЉЇђЅњ°£ґш‘Ў–уЅњќ™5.33—т/ha ±£ђ≤ЁµЎЌЅ»јЋЃЈ÷„оЇ√£®ЌЉ1-22£©°£Ћµ√ч ґ»“эЈҐƒЅ≤Ёµƒ≥ђ≤є≥•…ъ≥§£ђќђ≥÷≤ЁµЎЅЉЇ√µƒќп÷÷љбєєЇЌЌЅ»јћхЉю°£њ…“‘»ѕќ™ ґ»Ј≈ƒЅ”–јы”Џ≤ЁµЎї÷Єі£ђ «ќђї§≤ЁµЎљ°њµµƒ÷Ў“™ЌЊЊґ°£
≈ѓЉЊЈ≈ƒЅ£ђћ≤—т»’‘ц÷Ў“ї∞г‘Џ2.67—т/hm2ѕ¬„оЄя£®ЌЉ1-23£©°£2002ƒк£ђµЏ1¬÷Ј≈
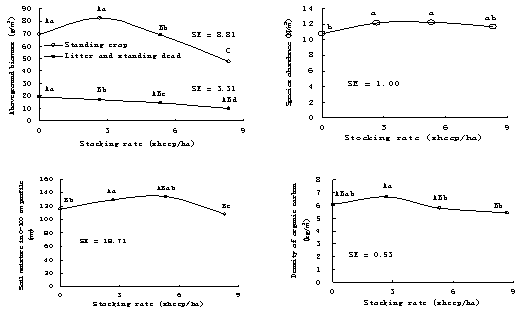
ЌЉ1-22ѕƒЉЊ¬÷ƒЅґ‘÷≤±їЇЌЌЅ»јµƒ”∞ѕм
„Ґ£ЇЌЅ»ј”–їъћЉќ™2004ЇЌ2005ƒкµƒ∆љЊщ÷µ£ђ∆д”аќ™2002-2005ƒк∆љЊщ÷µ


ЌЉ1-23≤їЌђЈ≈ƒЅ«њґ»Ј≈ƒЅћ≤—т…ъ≤ъЅ¶±дїѓ
„Ґ£ЇѕƒЉЊќ™2002ƒк-2006ƒк∆љЊщ÷µ£ђґђЉЊќ™2002-2005ƒк∆љЊщ÷µ
ƒЅ…х÷Ѕњ…“‘іпµљ176.7g/d£ђµЏ2¬÷ЇЌµЏ3¬÷Ј≈ƒЅ£ђ“≤њ…“‘‘Џ120g/d“‘…ѕ°£5.33—т/hm2ЇЌ8.67—т/haµƒћ≤—т»’‘ц÷ЎЈ÷±р‘Џ65°™167.5g/dЇЌ66.2g/d°™138.1g/d÷ЃЉд°£єэ»•5ƒк÷–£ђЈ≈ƒЅћ≤—тµ•÷їїоће‘ц÷ЎЋжЈ≈ƒЅ«њґ»‘ціу≥ љµµЌ«ч ∆°£‘Џ“їґ®‘Ў–уЅњЈґќІƒЏ£ђµ•ќї√жїэ≤ЁµЎЈ≈ƒЅћ≤—тµƒ∆љЊщ»’‘ц÷Ў≥ …ѕ…э«ч ∆°£“тќ™µ±Ј≈ƒЅ«њґ»≥ђєэ“їґ®ЈґќІЇу£ђ”…”Џ≤ЁµЎЋ•∞№£ђЉ“–у…ъ≤ъƒ№Ѕ¶Љ±Њзѕ¬љµ°£
“јЊЁ÷≤±ї”лЌЅ»јћЎ’ч£ђ»Јґ® ‘—й«ш≤ЁµЎ…ъ≥§ЉЊ£®6‘¬°™9‘¬£© “Ћ‘Ў–уЅњќ™3.6-4.1—тµ•ќї/ha°£Љ“–уµƒ…ъ≤ъЅ¶ћЎ’ч“≤њ…“‘„чќ™»Јґ® “ЋЈ≈ƒЅ«њґ»µƒ“јЊЁ£®ЌЉ1-23£©°£ЄщЊЁЌЅ»јЇЌ÷≤±їЋщ»Јґ®µƒ “ЋЈ≈ƒЅ«њґ»Ћд»ї‘Џ’вЄц÷µ”тЈґќІƒЏ£ђµЂ±»„оЄя÷µµЌ°£’вЈі”≥≥ц≤ЁµЎ…ъ≤ъ÷–£ђЊ≠Љ√–І“ж£®“‘ґѓќп…ъ≤ъќ™іъ±н£©”л…ъћђ–І“ж£®“‘÷≤±їЇЌЌЅ»ј„іњцќ™іъ±н£©µƒ√ђґ№£ђµЂґю’я «њ…µчЇЌµƒ°£…ъ≤ъ µЉщєэ≥ћ÷–£ђ “ЋµƒЈ≈ƒЅ«њґ»–и“™“јЊЁЌЅ°™≤Ё°™–у„џЇѕћЎ’ч»Јґ®°£±Њѕо—–Њњ“‘…ъћђ–І“жќ™÷ч£ђЉжєЋЊ≠Љ√–І“ж£ђ”√ЌЅ°™≤Ё°™–уѕаї•єЎѕµ»Јґ® “Ћ‘Ў–уЅњќ™4.3—т/ha°™5.6—т/ha£ђ≤ЁµЎµƒґѓќп…ъ≤ъƒ№Ѕ¶ќ™41.9APU/ha°™51.2APU/ha°£
ґђЉЊ¬÷ƒЅ ‘—й±н√ч£ђћ≤—т…ъ≤ъЅ¶≤їЌђ≥ћґ»ѕ¬љµ£®ЌЉ1-23£©£ђЈ≈ƒЅ‘љ«б£ђћ≤—тЄцће І÷Ў„о…ў°£і”–у≤ъ∆Ј≤ъЅњµƒљ«ґ»јіљ≤£ђґђЉЊ≤ї ЇѕЈ≈ƒЅ°£µЂ «£ђґ‘”Џ–у≤ъ∆Ј÷ Ѕњ…ѕїт’яїщі°–у»Їќђ≥÷£ђ»іЊя”–“їґ®µƒњ…–––‘°£»зєыљбЇѕ≤єЋ«£ђЌк»Ђњ…“‘±£÷§√а—тће÷Ў‘цЉ”£ђ…ъ≤ъ…ѕ“≤Њя”–њ…–––‘°£
µЂ «£ђі”–у»Ї‘љґђµƒљ«ґ»£ђ2002ƒкЇЌ2003ƒкћ≤—т‘Џ45dƒЏће÷ЎЈ÷±рѕ¬љµ5.1%-8.4%ЇЌ2.0%-4.0%°£Љ“–ућењц…–њ…љ” №£ђґш«“√ї”–≤єЋ«Ј—”√£ђЊ≠Љ√…ѕ“≤њ…––°£ґш«“Ј≈ƒЅ≤ЁµЎѕаґ‘”ЏќіЈ≈ƒЅµƒґ‘’’—щµЎ£ђ÷≤±їЇЌЌЅ»ј√ї”–ѕ‘÷ш±дїѓ£®p>0.05£©£ђ…ъћђ…ѕњ…––°£’вѕо—–Њњ£ђїє–иљбЇѕ≤єЋ«љш––£ђ“‘ћљЋчґђЉЊјы”√ћм»ї≤ЁµЎµƒ„оЉ—ЌЊЊґ°£
7.2.2 “ЋЈ≈ƒЅ ±Љдµƒ»Јґ®----¬÷ƒЅ≤ЁµЎљ°њµє№јнµƒ°∞г–ѕёЋЂ“т„”Ј®°±
ЄщЊЁƒЅ≤Ё…ъ≥§ґѓћђ£ђ“їЄц¬÷ƒЅ÷№∆Џњ…“‘Ј÷ќ™Ј≈ƒЅ∆ЏЇЌ‘ў…ъ≥§∆Џ£®ЌЉ1-24£©°£≤ЁµЎЈ≈ƒЅ≤ъ…ъ“їЄц÷≤ќп…ъјн…ъ≥§µƒµЌѕё£ђ’в «≤ЁµЎќђ≥÷њ…≥÷–шє¶ƒ№ЇЌљ°њµЋщƒ№≥– №µƒ„оµЌг–÷µ£ђґ‘”¶„≈≤ЁµЎ„оµЌЄяґ»°ҐЄ«ґ»їтѕ÷іжЅњµ»°£ƒЅ≤Ёµƒ‘ў…ъ≥§∆Џ”÷њ…“‘Ј÷ќ™µч’ы∆ЏЇЌї÷Єі∆Џ£®ЌЉ1-24£©°£ƒЅ≤Ё‘Џµч’ы∆Џ÷ч“™–ёЄі №Ћр…ъјнїъєє£ђ÷Ў–¬Ј÷≈д÷ь≤Ўќп÷ £ђќ™єЏ≤г÷Ўљ®ЇЌ‘ў…ъ≥§„ч±Ў“™µƒ…ъјн…ъ≥§„Љ±Є°£ƒЅ≤Ё‘Џї÷Єі∆Џ…ъјн…ъ≥§є¶ƒ№÷рљ•ї÷ЄіµљЈ≈ƒЅ«∞µƒљ°њµЋЃ∆љ£ђ≤Ґ≤ъ…ъ“їЄц…ъјн…ъ≥§µƒ…ѕѕё£®ЌЉ1-24£©£ђ’в «ƒЅ≤Ё…ъјн…ъ≥§їоґѓƒ№єїї÷Єіµƒ„оЄяг–÷µ£ђґ‘”¶„≈≤ЁµЎ„оіуѕ÷іжЅњїт÷ь≤ЁЅњµ»°£‘ЏЈ≈ƒЅ∆Џ£ђƒЅ≤Ёµƒ…ъјн…ъ≥§÷Є±кљ”љь”Џ’э≥£µƒ…ъ≥§µЌѕёг–÷µ£ђ»фЉћ–шЈ≈ƒЅ£ђ‘т≤ЁµЎƒ—“‘ї÷Єі£ђ“тіЋ£ђ…ъ≥§µЌѕёг–÷µњ…„чЈ≈ƒЅљб шµƒ÷Є±к°£ƒЅ≤Ё‘ў…ъ≥§∆Џ£ђƒЅ≤Ё…ъ≥§іпµљ…ъ≥§Єяѕёг–÷µ£ђ»ф≤ЁµЎ»‘≤їјы”√£ђЊЌїб‘м≥…„ ‘іјЋЈ—£ђґш«“≤Ё»Їљбєє±дїµ£ђ…ъ≥§Єяѕёг–÷µњ…„чќ™Ј≈ƒЅњ™ Љµƒ÷Є±к°£Ћщ“‘£ђЌ®єэ ‘—й»Јґ® “ЋµƒµЌѕёг–÷µЇЌЄяѕёг–÷µ£ђњ…“‘÷ЄµЉ≤ЁµЎµƒљ°њµє№јн£ђ’вЊЌ «≤ЁµЎљ°њµє№јнµƒЋЂ“т„”г–ѕёЈ®°£ґ‘”ЏЈ≈ƒЅ≤ЁµЎ£ђƒЅ≤Ё‘Џ∆чєў°ҐЄцће°Ґ÷÷»ЇЇЌ»Ї¬дµ»ЄчЄц„й÷ѓ≤гіќґЉіж‘ЏЅљЄц°£‘Џћм»ї≤ЁµЎ£ђ”…”ЏЉ“–уµƒ‘с ≥–‘ЇЌƒЅ≤ЁњєƒЅ–‘µƒ÷÷Љд≤о“м£ђƒЅ≤Ё÷÷÷ЃЉдµƒ…ъјн…ъ≥§±дїѓ≤їЌђ≤љ£ђЄчѕо÷Є±к±д“м¬ Ї№Єя£ђ“тіЋ»Јґ®ЋЂ“т„”µƒ»Їћег–ѕё÷µЄьЊя µЉщЉџ÷µ£ђ»з»Ї¬дЄ«ґ»°ҐµЎ…ѕѕ÷іжЅњ°Ґµт¬дќпѕ÷іжЅњµ»£ђґЉ «»Ї¬д…ъ≤ъЅ¶µƒ±нѕ÷–ќ љ°£
‘ЏѕƒЉЊЈ≈ƒЅµЎ£ђЄщЊЁг–ѕёЋЂ“т„”Ј®£ђ≤…”√ґа÷÷÷≤±ї÷Є±к£ђњ…“‘»Јґ®≤їЌђљµЋЃћЎ’чµƒƒкЈЁ£ђЈ≈ƒЅµЎЇќ ±њ™ ЉЈ≈ƒЅЇЌљб шЈ≈ƒЅ£®ЌЉ1-24£©£ђ Љ÷’±£≥÷Ј≈ƒЅ≤ЁµЎµƒљ°њµ„іћђ°£Є√ЈљЈ®‘Џ ‘—й«шЈ≈ƒЅѕµЌ≥÷–µ√µљ‘Ћ”√£ђ»°µ√ЅЉЇ√–Ієы°£



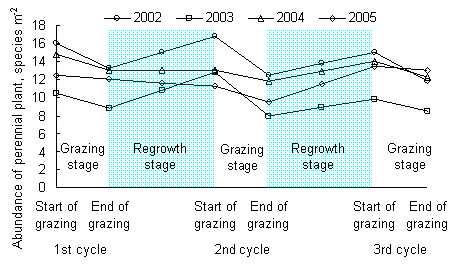
ЌЉ1-24 ≈ѓЉЊ¬÷ƒЅ∆ЏЉд£ђ≤ЁµЎ…ъќпЅњ”лќп÷÷ґѓћђ
7.3 °∞ЌЋЄыµЎЈв”э°±µƒ„‘»її÷ЄіЉЉ х
‘Џ—–Њњ«ш…и÷√“—ї÷Єі1ƒк°Ґ2ƒк°Ґ4ƒкЇЌ7ƒкµƒЌЋЄыµЎ£ђ“‘ћм»ї≤ЁµЎ„чќ™ґ‘’’£ђ±»љѕ÷≤±ї”лЌЅ»јµƒї÷Єіљш≥ћ°£
ї∆ЌЅЄя‘≠ЌЋЄыµЎї÷Єіљш≥ћ÷–£ђ÷≤±їі”…≥≈”≈»Ї¬дѕт“‘іпќЏјпЇъ÷¶„”ќ™”≈ ∆÷÷µƒ»Ї¬д—Ёћж°£ќп÷÷µƒ‘цЉ”Ћўґ»£ђ≥х∆Џњм£ђЇу∆Џ¬э°£ЌЋЄыЇуї÷Єі≥х∆Џµƒ–¬‘цќп÷÷÷ч“™ «…≥≈оЇЌ∞„≤Ё£ђ÷–«∞∆Џ÷ч“™ «“р≥¬Ёп£ђЋь√« «»Ї¬дµƒіќ”≈ ∆÷÷£ї÷–Їу∆Џ–¬‘цќп÷÷÷ч“™ «ґаƒк…ъґєњ∆÷≤ќп£ђ÷ї «»Ї¬дµƒ∞й…ъ÷÷°£»Ї¬д÷–1ƒк…ъЇЌґаƒк…ъ÷≤ќпµЎ…ѕ…ъќпЅњЈ÷±р“‘8.8%µƒƒк∆љЊщЋў¬ Љх…ўїт‘цЉ”£ђґєњ∆÷≤ќпµЎ…ѕ…ъќпЅњ±»јэ…ѕ…э£®ЌЉ1-25£©°£ЌЋЄыµЎї÷Єієэ≥ћ÷–£ђ…ѕ≤гЌЅ»ј÷–»Ї¬дµЎѕ¬…ъќпЅњ±»јэ”л±дїѓЈщґ»‘цЉ”£ђ±нѕ÷≥цЄщѕµ«≥≤гїѓ«ч ∆°£…≥≈о÷÷»ЇЇЌіпќЏјпЇъ÷¶„”÷÷»Ї÷кЄя°ҐєЏЈщ÷±Њґ°ҐЈ÷÷¶ эЇЌ…ъ÷≥÷¶ эЋжї÷Єі ±Љд≥ ѕаЈіµƒ±дїѓ«ч ∆°£ЌЋЄыµЎї÷Єі—Ёћж1°™7ƒкЌЅ»ј’≥Ѕ£ЇЌЈџЅ£Љх…ў£ђ…∞Ѕ£‘цЉ”£ђ7ƒкЇу±дїѓ«ч ∆ѕаЈі°£ЌЋЄыµЎ0-100cmЌЅ≤гЋЃЈ÷ЇђЅњ‘Џї÷Єі∆ЏЉд≥ ÷рљ•…ѕ…э«ч ∆£ђ÷–∆Џ‘цЈщѕ‘÷ш°£»ЂNЇЌЋў–ІN‘Џї÷Єі«∞∆ЏЉх…ў£ђЇу∆Џ‘цЉ”°£Ў„Єојы”√ѕ¬£ђЌЋЄыµЎї÷Єієэ≥ћ÷–ЌЅ»ј»ЂP°ҐЋў–ІPЇЌ”–їъC≥÷–шЋ•љя£®ЌЉ1-25£©°£
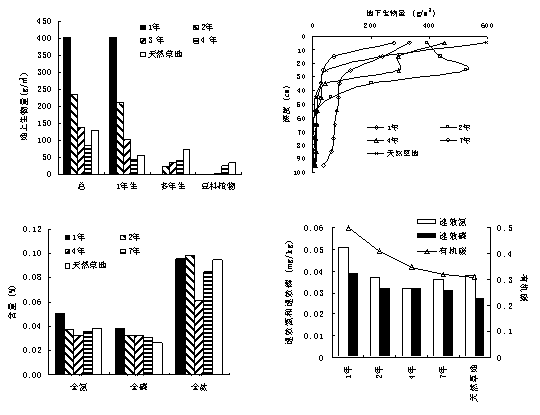
ЌЉ1-25 ЌЋЄыµЎї÷Єіљш≥ћ÷–÷≤±ї”лЌЅ»јћЎ’ч
ЄщЊЁ÷÷»Ї°Ґ»Ї¬дЇЌЌЅ»јћЎ’чѕаґ‘”Џћм»ї≤ЁµЎµƒ±дїѓЋўґ»£ђЌЋЄыµЎї÷Єіµљµ±«∞ќ»ґ®µƒ»Ї¬дЋщ–и ±ЉдЈ÷±рќ™£ђ”≈ ∆÷÷»Ї8°™9ƒк£ђ»Ї¬д9°™11ƒк£ђЌЅ»ј11°™12ƒк£®±н1-27£©°£≥§∆ЏЄы„чЄƒ…∆ЅЋЌЅ»јљбєє”лЈ Ѕ¶£ђ“тіЋ‘ЏЌЋЄыµЎ∆ъЄы‘з∆Џ£ђ»Ї¬дЊя”–љѕЄяµƒ…ъќпЅњ£ї∆ъЄыµЎµƒї÷Єі—ЁћжЋўґ»“≤±»»Ї¬дµƒ‘≠…ъ—Ёћжњмµ√ґа£ђ≤Ґ«“∆ъЄыµЎі”“їњ™ ЉЊЌЊя”–љѕЄяµƒ…ъћђЈјї§ƒ№Ѕ¶°£Ћщ“‘£ђ‘Џ—–Њњ«ш”т£ђґа эЄыµЎ”√∆ъЄыЈв”эµƒЈљ љњ™’є…ъћђї÷Єі «њ…––µƒ°£
±н1-27 ЌЋЄыµЎµƒї÷Єі ±Љд
|
|
÷Є±к |
ї÷Єі ±Љд £®ƒк£© |
|
÷÷»Ї |
…≥≈о |
8.4°ј0.2 |
|
іпќЏјпЇъ÷¶„” |
8.9°ј0.4 |
|
∆љЊщ |
8.7°ј0.3 |
|
»Ї¬д |
»Ї¬дѕаЋ∆–‘ |
9.5 |
|
ќп÷÷ЈбЄїґ» |
10.4 |
|
µЎ…ѕѕ÷іжЅњ |
12.6 |
|
ґаƒк…ъ÷≤ќп/1ƒк…ъ÷≤ќпµЎ…ѕ…ъќпЅњ±»јэ |
8.6 |
|
∆љЊщ |
10.3°ј0.9 |
|
ЌЅ»ј
|
ЌЅ»јЅ£Њґ£®’≥Ѕ£°ҐЈџЅ£ЇЌ…≥Ѕ££© |
11.4 |
|
ЌЅ»јЈ Ѕ¶£®»Ђµ™ЇЌЋў–Іµ™£© |
11.9 |
|
∆љЊщ |
11.6°ј0.4 |
8. ЌЋїѓ°Ґ…≥їѓ≤ЁµЎ÷≤±її÷ЄіЉЉ х—–Њњ
Ѕ÷ќчѕЎ”–љь10ЌтєЂ«кµƒћм»ї≤ЁµЎ£ђ”…”Џ≥§∆Џ≥ђ‘ЎєэƒЅЇЌ√§ƒњњ™њ—£ђ≤ЁµЎЌЋїѓ°Ґ…≥їѓ»’«ч—ѕ÷Ў£ђЌЋїѓ°Ґ…≥їѓ≤ЁµЎ√жїэ’Љњ…јы”√≤ЁµЎ√жїэ85%“‘…ѕ°£ќ™ЅЋї÷Єі≤ЁµЎ÷≤±ї£ђћбЄя≤ЁµЎ…ъ≤ъЅ¶£ђќ“√«њ™’єЅЋЌЋїѓ°Ґ…≥їѓћм»ї≤ЁµЎ÷≤±її÷ЄіЉЉ х—–Њњ”л ЊЈґЌ∆єг°£‘Џ…о»л—–Њњ»ЂѕЎ≤ЁµЎ…ъћђѕµЌ≥ №Ћрїъјнµƒїщі°…ѕ£ђ‘Ћ”√ќІЈвљыƒЅ°Ґ–ЁƒЅµ»„‘»її÷ЄіЇЌ—°‘с ”¶–‘«њµƒƒЅ≤Ёљш––≤є≤•µ»ЉЉ хіл ©£ђє≤ЄƒЅЉ…≥їѓ°ҐЌЋїѓ°Ґ—ќЉоїѓ≤ЁµЎ4100hm2£ђ є÷≤±їЄ«ґ»і”‘≠јі≤ї„г30%ћбЄяµљ80%“‘…ѕ£ђƒЅ≤Ё≤ъЅњі”434.69°Ђ750.01kg/hm2ћбЄяµљ1464.04°Ђ2850.12kg/hm2£ђµЎѕ¬…ъќпЅњ“≤√чѕ‘‘цЉ”£ђ≤ЁµЎќІЈв4ƒкЇу£ђµЎѕ¬…ъќпЅњ”…‘≠јіµƒ4.41g‘цЉ”µљ28.49g(≥§°Ѕњн°Ѕ…оґ»=20cm°Ѕ20cm°Ѕ30cm)°£…≥їѓ°ҐЌЋїѓ≤ЁµЎ÷≤±їµ√µљ√чѕ‘µƒї÷Єі£ђ»Ї¬дљбєєЈҐ…ъѕ‘÷ш±дїѓ£ђі”ґшґф÷∆ЅЋ≤ЁµЎЌЋїѓµƒћђ ∆°£
—°‘с«бґ»їт÷–ґ»ЌЋїѓ≤ЁµЎљш––ќІЈвљыƒЅ£ђ4ƒкЇу£ђ≤ЁµЎ÷≤±їµ√µљ√чѕ‘µƒї÷Єі£ђ»Ї¬дљбєєЈҐ…ъ√чѕ‘±дїѓ£ђ÷≤ќп÷÷ја‘цґа£ђ”…‘≠јіµƒ ЃЉЄ÷÷‘цЉ”µљЉЄ Ѓ÷÷£®±н1-28£©°£
±н1-28°°÷≤ќп»Ї¬дљбєєћЎ–‘
|
—щµЎ
√ы |
јы”√
Јљ љ |
”≈ ∆
÷≤ ќп |
÷≤ ќп ÷÷ ја |
|
іуЋЃ≤§¬ёƒЅ≥°
|
»ЂƒкЈ≈ƒЅ |
≤Џ“ю„”≤Ё°Ґ’л√©°Ґ
ј«ґЊ |
ј«ґЊ°Ґ≤Ё№њѕг°Ґїр»ё≤Ё°Ґ≤Џ“ю„”≤Ё°Ґіу’л√©°Ґ—т≤Ё°Ґ≤ЏЋ’°Ґ±в–оґє°ҐќѓЅк≤Ћ°Ґћ∆Ћ…≤Ё°ҐѕЄ“ґі–°Ґ≤сЇъ°ҐЅ—“ґЁп°ҐЈјЈз°ҐµЎ”№°ҐѕЄ“ґр∞ќ≤ |
|
∞лƒкљыƒЅ(ґђЉЊЈ≈ƒЅ) |
ѕя“ґЊ’°Ґ
іу’л√©°Ґ
Љв“ґЇъ÷¶„” |
ѕя“ґЊ’°Ґіу’л√©°ҐЉв“ґЇъ÷¶„”°Ґїр»ё≤Ё°Ґ≤Ё№њѕг°Ґј«ґЊ°ҐµЎ”№°ҐЇµ¬у∆њ°Ґ∞Ґґыћ©єЈЌёї®°ҐЅ—“ґЁп°Ґ≤ЏЋ’°Ґћ¶≤Ё°Ґ≤Џ“ю„”≤Ё°Ґ«Ґ≤Ё°Ґ…дЄ…р∞ќ≤°Ґї∆№ќ°ҐЈзЅе≤Ё°ҐґюЅ—ќЃЅк≤Ћ°Ґ≤сЇъ°ҐЉђґє°Ґ…љ“∞Ќгґє°Ґ±в–оґє°ҐѕЄ“ґі–°Ґ ѓ÷с°Ґ¬йї®ЌЈ°ҐѕЄ“ґр∞ќ≤°Ґ«Ў№і°Ґ–«√ЂќЃЅк≤Ћ°Ґё§°Ґ–±Њ•ї∆№ќ°ҐЌяЋ…°ҐЈјЈз°Ґґћї®’л√©°Ґ≤ЁƒЊйЎ„ії∆№ќ°Ґ≥µ÷б≤Ё°ҐіћЋлёЉ°ҐЊ£љж°Ґ≈о„”≤Ћ°ҐЌЅ»э∆я°ҐЌбЌЈ≤Ћ°ҐјдЁп |
|
–¬≥«„”’т |
»ЂƒкЈ≈ƒЅ |
∞ўјпѕг°ҐјдЁп°ҐєЈќ≤≤Ё°Ґ |
іпќЏјпЇъ÷¶„”°Ґ∞ўјпѕг°ҐјдЁп°ҐЉђґє°ҐµЎљх°Ґ∞Ґґыћ©єЈї®°ҐєЈќ≤≤Ё°ҐїҐќ≤≤Ё |
|
іт≤Ё≥° |
іпќЏјпЇъ÷¶„”°Ґ—т≤Ё°Ґ
÷–ї™“ю„”≤Ё |
іпќЏјпЇъ÷¶„”°Ґ∞ўјпѕг°ҐјдЁп°ҐЉђґє°Ґ÷–ї™“ю„”≤Ё°Ґ”р√©°Ґ—т≤Ё°Ґіу“ґ’Ѕ°Ґ≤ЁƒЊйЎ„ії∆№ќ°Ґ≤Ё№њѕг°ҐќЃЅк≤Ћ°Ґ÷™ƒЄ°ҐѕЄ“ґі–°Ґ≤Ё¬йї∆°Ґ…дЄ…р∞ќ≤
|
|
ЋЂЊЃµкѕз |
»ЂƒкЈ≈ƒЅ |
“їƒк…ъ‘”≤Ё |
іпќЏјпЇъ÷¶„”°ҐјдЁп°ҐЉђґє°Ґ≤Џ“ґї∆№ќ°ҐѕЄ“ґр∞ќ≤°ҐµЎљх°Ґ÷є—™¬нћ∆°Ґіз≤Ёћ¶°ҐєЈќ≤≤Ё |
|
»ЂƒкљыƒЅ |
—т≤Ё°Ґ
±щ≤Ё°Ґ
іпќЏјпЇъ÷¶„” |
—т≤Ё°Ґ±щ≤Ё°ҐіпќЏјпЇъ÷¶„”°Ґ≤Џ“ю„”≤Ё°Ґї≠√Љ≤Ё°Ґ∞„≤Ё°ҐќЃЅк≤Ћ°ҐјдЁп°ҐѕЄ“ґр∞ќ≤°ҐµЎљх°Ґіз≤Ёћ¶°Ґ±в–оґє°Ґјµ≤Ё°Ґќч∞Ўјы—«”р√©°Ґ∞ђЁп°ҐјдЁп°ҐµЎ…’єѕ°Ґ÷н√Ђ≤Ћ°ҐїҐќ≤≤Ё°ҐєЈќ≤≤Ё |
≤Ё≤гЄяґ»”…‘≠јіµƒ≤ї„г10cmћбЄяµљ20°Ђ35cm £ђ”≈ЅЉґєњ∆ƒЅ≤ЁЇЌЇћ±Њњ∆ƒЅ≤Ё√чѕ‘‘цґа£ї≤ЁµЎ≤ъ≤ЁЅњµ√µљѕ‘÷шµƒћбЄя£ђ∆д÷–ґєњ∆ЇЌЇћ±ЊƒЅ≤ЁЋщ’Љ±»јэ‘цЉ”(±н1-29)°£
±н1-29ћм»ї≤ЁµЎµЎ…ѕ…ъќпЅњ£® kg/hm2£©
|
—щµЎ√ы |
јы”√Јљ љ |
„№…ъќпЅњ |
ґєњ∆ƒЅ≤Ё |
Їћ±Њњ∆
ƒЅ≤Ё |
‘”ја≤Ё |
|
іуЋЃ≤§¬ёƒЅ≥°
іуЋЃ≤§¬ёƒЅ≥°
–¬≥«„”’т
–¬≥«„”’т
ЋЂЊЃµкѕз
ЋЂЊЃµкѕз
–Ћ¬°„ѓѕз
–Ћ¬°„ѓѕз |
»ЂƒкЈ≈ƒЅ
∞лƒкљыƒЅ
»ЂƒкЈ≈ƒЅ
іт≤Ё≥°
»ЂƒкЈ≈ƒЅ
»ЂƒкљыƒЅ
Ј…≤•Јв”эіт≤Ё
«≥Єы≤є≤•іт≤Ё |
426.21
1874.24
436.69
902.25
119.66
687.58
1112.86
1464.04 |
138.97
53.65
214.61
155.18
914.96
640.32 |
200.20
321.06
294.50
69.06
41.28
236.17
179.89
427.52 |
226.01
1414.21
88.54
618.61
74.38
296.23
18.01
396.20 |
…≥їѓ°ҐЌЋїѓ≤ЁµЎ‘ЏљыƒЅ°ҐЈв”эµƒєэ≥ћ÷–£ђ≥эµЎ…ѕ≤њЈ÷÷≤±їµ√µљї÷ЄіЌв£ђµЎѕ¬…ъќпЅњ“≤ЈҐ…ъЅЋ√чѕ‘µƒ±дїѓ(±н1-30)°£
±н1-30 ћм»ї≤ЁµЎµЎѕ¬…ъќпЅњ (g°Ґcm)
|
—щµЎ√ы |
јы”√Јљ љ |
„№ЄщЅњ |
Єщ≤г |
|
0°Ђ10 |
10°Ђ20 |
20°Ђ30 |
|
іуЋЃ≤§¬ёƒЅ≥°
іуЋЃ≤§¬ёƒЅ≥°
–¬≥«„”’т
–¬≥«„”’т
ЋЂЊЃµкѕз
ЋЂЊЃµкѕз |
»ЂƒкЈ≈ƒЅ
∞лƒкљыƒЅ
»ЂƒкЈ≈ƒЅ
іт≤Ё≥°
»ЂƒкЈ≈ƒЅ
»ЂƒкљыƒЅ |
19.84
32.15
4.41
28.49
1.99
12.74 |
17.21
25.45
3.95
25.75
0.82
9.98 |
2.03
3.95
0.24
1.67
0.74
1.85 |
0.60
2.75
0.22
1.07
0.43
0.91 |
|
—°‘сЄ«ґ»µЌ”Џ30%µƒ÷Ўґ»ЌЋїѓ≤ЁµЎЇЌ…≥їѓ≤ЁµЎљш––«≥Єы≤є≤•ЇЌ»Ћє§ƒ£ƒвЈ…≤•£ђ»°µ√ЅЋљѕЇ√µƒ–Ієы£ђ≤є≤•≤Ё÷÷—°‘с ”¶–‘«њ°ҐƒЌі÷Ј≈є№јнµƒЉв“ґЇъ÷¶„”°ҐіпќЏјпЇъ÷¶„”°Ґ±вЋл±щ≤ЁЇЌ√…є≈±щ≤Ёљш––ЌЋїѓ≤ЁµЎ«≥Єы≤є≤•£ђ≤ъ≤ЁЅњњ…іп1464.04kg/hm2£ђ∆д÷–£ђґєњ∆ƒЅ≤ЁЇЌЇћ±Њњ∆ƒЅ≤ъЅњЈ÷±ріпµљ640.32 kg/hm2ЇЌ427.52 kg/hm2£ї…≥їѓ≤ЁµЎ»Ћє§ƒ£ƒвЈ…≤•≤ъ≤ЁЅњњ…іп1112.86 kg/hm2£ђ∆д÷–ґєњ∆ƒЅ≤ЁЇЌЇћ±Њњ∆ƒЅ≤Ё≤ъЅњЈ÷±ріпµљ914.96 kg/hm2ЇЌ179.89 kg/hm2°£«≥Єы≤є≤•ґ‘‘≠ Љ÷≤±їµƒ∆∆їµ–‘љѕіу£ђЄƒЅЉЇу1°Ђ2ƒк≤ЁµЎ…ъ≤ъЅ¶љѕµЌ£ђµЂµЏ3ƒк“‘Їу≤ъ≤ЁЅњЊз‘ц£ђќ™Јј÷єЌЅµЎ…≥їѓ∆рЉы£ђЄ√ЉЉ х‘Џ≤…”√ ±”¶…ч÷Ў°£»Ћє§ƒ£ƒвЈ…≤•ґ‘≤ЁµЎ‘≠ Љ÷≤±ї≤їїб‘м≥…∆∆їµ£ђ “Ћ‘Џ…≥їѓ≤ЁµЎ…ѕ µ ©£ђ’в÷÷ЉЉ х≤ў„чЉтµ•£ђ±г”Џ»Ї÷Џ’∆ќ’£ђ «ЄƒЅЉ…≥їѓ≤ЁµЎ––÷Ѓ”––ІµƒЇ√ЉЉ х°£
Ќ®єэћм»ї≤ЁµЎ÷≤±її÷ЄіЉЉ х—–Њњ£ђ»°µ√ЅЋ»зѕ¬љбєы£ЇҐў‘Џ«бґ»їт÷–ґ»ЌЋїѓ≤ЁµЎ…ѕ£ђ≤…”√љыƒЅЈв”эЉЉ х£ђњ…»°µ√љѕЇ√µƒ–Ієы£ђЊ≠єэ4ƒкµƒљыƒЅЈв”э£ђЌЋїѓ°Ґ…≥їѓ≤ЁµЎ÷≤±їµ√µљ√чѕ‘ї÷Єі£ђ»Ї¬дЈҐ’єљш’є—Ёћж£їҐЏ‘ЏЌЋїѓ≤ЁµЎ—Ё±дєэ≥ћ÷–£ђ»Ї¬д÷–µƒ”≈ ∆÷÷»Їіж‘Џ„≈–≠Ќђ±дїѓ£їҐџ”лЌЋїѓ≤ЁµЎѕа±»£ђ…≥їѓ≤ЁµЎ„‘»їЈв”эЋўґ»љѕ¬э£ђћЎ±р «µЎѕ¬≤њЈ÷£їҐ№»Ћє§ƒ£ƒвЈ…≤•ЉЉ х≤ў„чЉтµ•£ђ‘Џ÷Ўґ»ЌЋїѓµƒ∞лЅчґѓ…≥µЎ≤…”√Є√ЉЉ х£ђƒ№—ЄЋў‘цЉ”÷≤±ї£ђ–ќ≥…“‘Ј…≤•”≈ЅЉƒЅ≤Ёќ™”≈ ∆µƒ÷≤ќп»Ї¬д£ђ‘ЏЈв”эћхЉюѕ¬ЅљƒкЉіњ…»°µ√ѕ‘÷ш≥…–І°£
9. ≤ЁµЎ…ъћђѕµЌ≥Љџ÷µ∆јєјЉ∞∆дґѓћђƒ£ƒв
±Њ—–Њњ «‘Џґ‘µд–Ќ«ш”тµч≤й—–Њњµƒїщі°…ѕ£ђ‘Ћ”√…ъћђЊ≠Љ√—І°Ґ–уƒЅ—І°Ґ≤Ё“µњ∆—Іµ»ѕаєЎ—Іњ∆µƒ÷™ ґ£ђ≤…”√єжЈґ—–Њњ”л µ÷§—–ЊњѕаљбЇѕ£ђґ®–‘Ј÷ќц”лґ®ЅњЈ÷ќцѕаљбЇѕµƒЈљЈ®£ђљбЇѕќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥µƒ µЉ £ђґ‘ќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥µƒЉџ÷µЉ∞∆дє№јн≤я¬‘љш––ЅЋ—–Њњ£ђ’є ЊЅЋ—–ЊњњтЉ№µƒјн¬џЉџ÷µЇЌ”¶”√Љџ÷µ£ђћљћ÷ЅЋ…ъћђѕµЌ≥Љџ÷µ∆јєјµƒјн¬џ”лЉЉ х£ђќ™≤ЁµЎ…ъћђѕµЌ≥µƒє№јн°Ґї÷Єі”л±£ї§µƒњ∆—ІЊц≤яћбє©ЅЋјн¬џ“јЊЁ°£
9.1. ≤ЁµЎ…ъћђѕµЌ≥Љџ÷µµƒƒ№÷µ∆јєјњтЉ№—–Њњ
’лґ‘ƒњ«∞…ъћђѕµЌ≥Љџ÷µ∆јєјњтЉ№іж‘Џµƒ÷ч“™ќ ћв£Ї1£©°ҐЉџ÷µ∆јєјµƒјн¬џ…–і¶‘ЏћљЋчєэ≥ћ÷–£ђќіљЂ∆дƒ…»лЌґ„ Ј÷ќц”лЊц≤яєэ≥ћ£ї2£©°ҐЈ÷јаЄі‘”£ђЊ≤ћђЈ÷ќцґа£ђґѓћђЈ÷ќц…ў£ђґ®Љџіш”–√чѕ‘µƒ÷чєџ…Ђ≤ °£”¶”√ƒ№÷µјн¬џ”л…ъћђ—І°ҐЊ≠Љ√—Іјн¬џ”–їъљбЇѕ£ђћб≥цЅЋ≤ЁµЎ…ъћђѕµЌ≥Љџ÷µµƒƒ№÷µ∆јєјњтЉ№£®ЌЉ1-26£©°£
9.1.1…ъћђ„ ≤ъµƒ÷ч“™ƒЏ»Ё
1)…ъћђѕµЌ≥ЅҐµЎ∆љћ®µƒЉџ÷µ£ї2)…ъћђѕµЌ≥÷ьіжЋЃЉ∞‘™ЋЎµƒЉџ÷µ£ї3)ЌЅ»јƒаћњЉ∞”–їъ÷ Љџ÷µ£ї4) їоµƒ…ъќпЅњµƒЉџ÷µ£ї5)ћЎ”–ќп÷÷µƒїщ“т–≈ѕҐЉџ÷µ°£»зєыƒ≥÷÷ќп÷÷≤ҐЈ«±ЊµЎґј”–£ђњ…ЄщЊЁ∆д’Љ»Ђ«тЈ÷≤Љ√жїэµƒ±»јэљш––њпЋг°£
9.1.2 ≤ЁµЎ…ъћђѕµЌ≥є¶ƒ№
…ъћђѕµЌ≥Јюќсє¶ƒ№µƒ÷ч“™ƒЏ»Ё”–…ъћђє¶ƒ№°ҐЊ≠Љ√є¶ƒ№ЇЌ…зїбє¶ƒ№°£
9.1.2.1 ≤ЁµЎ…ъћђѕµЌ≥µƒ…ъћђє¶ƒ№
≤ЁµЎ…ъћђѕµЌ≥ «“їЄц”…∆шЇт°ҐµЎ–ќ°ҐЌЅ»ј°Ґ»’єв°Ґњ’∆ш°ҐЋЃЈ÷°ҐµЎѕ¬„ ‘і°Ґ÷≤ќп°Ґ
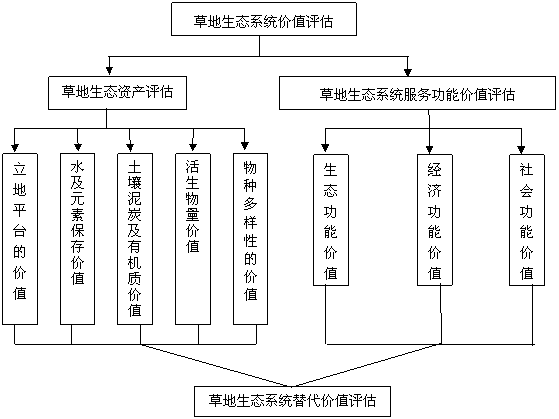
ЌЉ1-26 ≤ЁµЎ…ъћђѕµЌ≥Љџ÷µ∆јєјњтЉ№Љ∞∆дµƒ”–їъєє≥…
ґѓќп“‘Љ∞ќҐ…ъќпµ»„й≥…µƒ…ъћђѕµЌ≥£ђ∆д…ъћђЈюќсє¶ƒ№ «≤ЁµЎ…ъћђѕµЌ≥”л…ъћђєэ≥ћЋщ–ќ≥…Љ∞Ћщќђ≥÷µƒ»Ћјајµ“‘…ъіжµƒ„‘»їїЈЊ≥ћхЉюЇЌ–І”√°£÷ч“™±нѕ÷ «£ЇµчљЏ∆шЇт°ҐЄƒЅЉЌЅ»ј°Ґ±£≥÷ЋЃЌЅ°ҐЈјЈзєћ…≥°ҐЇ≠—шЋЃ‘і°Ґ√јїѓїЈЊ≥°ҐЊїїѓњ’∆ш°ҐЈј÷є‘л“фµ»ґа÷÷…ъћђЈюќсє¶ƒ№°£Ќ®єэЇѕјнµЎЈҐї”≤ЁµЎ…ъћђѕµЌ≥µƒ…ъћђє¶ƒ№£ђƒ№ є»Ћја…ъїо‘Џ“їЄц«еља”≈√јµƒ…ъїоїЈЊ≥ЇЌ…ъћђїЈЊ≥÷Ѓ÷–£ђљѕЇ√µЎЈј÷ќЄч÷÷ќџ»Њґ‘»Ћјаµƒњ…ƒ№Є…»≈ЇЌќ£Ї¶£ђ“‘Љ∞»ЋњЏµƒ’э≥£‘ў…ъ≤ъЇЌЄч÷÷≤ъ“µµƒЇѕјн‘ў…ъ≤ъ°£
9.1.2.2 ≤ЁµЎ…ъћђѕµЌ≥µƒЊ≠Љ√є¶ƒ№
≤ЁµЎµƒЊ≠Љ√є¶ƒ№ «≤ЁµЎЊ≠Љ√ѕµЌ≥‘Џ…ъћђѕµЌ≥љш––ќп÷ —≠їЈ°Ґƒ№Ѕњ„™їїЇЌ–≈ѕҐіЂµЁµƒЌђ ±£ђЄчјаЊ≠Љ√“™ЋЎµƒЌґ»лЇЌ≤ъ≥ц–ќ≥…ЅЋ¬ъ„г»Ћја≤їЌђ–и«уµƒЄч÷÷”––ќЇЌќё–ќµƒ÷–Љд≤ъ∆ЈЇЌ„о÷’≤ъ∆Ј£ђ‘ўЌ®єэ”––ќЇЌ“ю–ќ –≥°µƒљїїї£ђћбє©Єш±ЊµЎ«ш…ъ≤ъѕыЈ—°ҐЊ”√сѕыЈ—ЇЌЊ≠єэєъƒЏЌв√≥“„¬ъ„гєъƒЏЌв –≥°–и«уµƒ÷оє¶ƒ№µƒ„№≥∆°£∞ьј®≤ЁµЎ≤ъ∆Ј±Њ…нµƒЊ≠Љ√є¶ƒ№£ђїє”–Єƒ…∆їЈЊ≥°Ґ√јїѓ…з«шіўљш∆дЋь––“µ£ђ»з–уƒЅ“µ°Ґ¬√”ќ“µЇЌЈњµЎ≤ъ“µ‘ц÷µµƒє¶ƒ№°£
Ќ®єэЈҐї”≤ЁµЎ…ъћђѕµЌ≥µƒЊ≠Љ√є¶ƒ№£ђ‘Џ„‘»їќп÷ —≠їЈ÷––ќ≥…ЅЋќп÷ Ѕч°Ґƒ№ЅњЅч°Ґ–≈ѕҐЅчµƒЌђ ±£ђ“≤–ќ≥…ЅЋЊ≠Љ√Ѕч°£Љі“їЈљ√ж–ќ≥…ЅЋЄч÷÷ƒ№¬ъ„г»Ћја–и«уµƒЊ≠Љ√ќп÷ Љі є”√Љџ÷µ£ђ„чќ™’в≤њЈ÷Њ≠Љ√≤ъ∆Јµƒќп÷ ≥–µ£’я£їЅн“їЈљ√ж£ђ‘Џ’в≤њЈ÷Њ≠Љ√≤ъ∆ЈµƒЌґ»л≤ъ≥цєэ≥ћ÷–”÷≤їґѕµЎ„™“∆Љџ÷µ°Ґіі‘мЉџ÷µЇЌ‘ц÷µЉџ÷µ£ђЊ≠єэљїїї°ҐЈ÷≈д£ђ„о÷’љш»лѕыЈ—£ђ“‘ µѕ÷∆дЉџ÷µ°£і”ґш“≤ є„‘»їƒ№ЅњЅч”лЊ≠Љ√ƒ№ЅњЅч≤Ґіж£ђ є…ъћђѕµЌ≥ƒЏµƒ„‘»ї–≈ѕҐЅч”лЊ≠Љ√ѕµЌ≥µƒ…ъ≤ъ–≈ѕҐ°Ґ –≥°–≈ѕҐ°ҐѕыЈ—–≈ѕҐ≤Ґіж£ї≤Ґ є≤ЁµЎ…ћ∆ЈЇЌ≤ЁµЎЈюќс‘Џ…ъ≤ъЇЌЅчЌ®÷– µѕ÷Љџ÷µ‘ц÷µ°£
9.1.2.3 ≤ЁµЎ…ъћђѕµЌ≥µƒ…зїбє¶ƒ№
‘Џ≤ЁµЎ÷≤±їµƒ÷Џґає¶ƒ№÷–£ђ√чѕ‘±ї»Ћ√« µЉ є”√µƒє¶ƒ№ «¬ъ„гЊ”√с”йј÷°Ґќƒїѓљћ”э°Ґњ∆—І∆’Љ∞µ»ЊЂ…с…ъїоµƒ–и“™°£Ћж„≈…зїб”л»Ћјаµƒљш≤љЇЌЈҐ’є£ђЊ”√сґ‘…ъїоїЈЊ≥µƒ–и«у‘Џ≤їґѕћбЄя£ђ–Ёѕ–µƒ“в ґ‘Џ≤їґѕЉ”«њ°£њ…“‘Ћµ£ђ≤ЁµЎ…ъћђѕµЌ≥“—Њ≠≥…ќ™»Ћ√«…ъїо÷–µƒ“їЄц÷Ў“™≥…Ј÷£ђЊя”–÷Ў“™µƒ…зїбє¶ƒ№°£∆д…зїбє¶ƒ№÷ч“™±нѕ÷ќ™£Ї¬ъ„г…зїбЄчЈљ√ж–и«у£ђіўљшЊЂ…сќƒ√чљ®…и£їЄƒ…∆…ъ≤ъ…ъїоїЈЊ≥£ђ‘цљш»Ћ√«љ°њµ£їЄƒ…∆Ќґ„ їЈЊ≥£ђќь“эљ®…и„ љр£їћбє©ЅЉЇ√µƒ–ЁѕҐїЈЊ≥£ђїЇљв»Ћ√«–ƒјн—єЅ¶µ»°£
Ќ®єэ…ѕ цґ‘≤ЁµЎ÷≤±їѕµЌ≥µƒћЎµгЉ∞∆дљбєєµƒЈ÷ќц£ђњ…“‘ЈҐѕ÷£ђ≤ЁµЎѕµЌ≥ «“їЄцґа‘™µƒЄіЇѕѕµЌ≥£ђ“тіЋ£ђ≤ЁµЎ…ъћђѕµЌ≥±Ў»їЊя”–…ъћђ°ҐЊ≠Љ√”л…зїб»ЂЈљќїµƒЈюќсє¶ƒ№°£
≤ЁµЎ…ъћђѕµЌ≥ «“їЄцњ™Ј≈µƒѕµЌ≥£ђ”лЌвљзіж‘Џ„≈ƒ№Ѕњќп÷ µƒљїїї£ђ“тґш∆дЊя”–„‘ќ“‘ц÷µµƒє¶ƒ№°£ќ™ЅЋЅЋљвє¶ƒ№µƒґѓћђ£ђќ“√«њ…“‘—°‘с÷ч“™µƒ÷Є±кљш––ґѓћђƒ£ƒв°£»зґ‘”Џ≤ЁµЎ÷≤±ї£ђќ“√«њ…“‘—°‘сїо…ъќпЅњ°ҐЊї…ъ≤ъЅ¶°ҐќьћЉЈ≈—хє¶ƒ№µ»єЎЉь÷Є±кґ‘…ъћђ„ ≤ъ°ҐЈюќсє¶ƒ№µƒєЎЉь÷Є±кљш––„џЇѕЈ÷ќц°£»ф“™ЅЋљвЄьґає¶ƒ№µƒґѓћђ£ђњ…‘цЉ”Ј÷ “ƒ£–ЌµƒƒЏ»Ё°£
9.2 ≤ЁµЎ…ъћђѕµЌ≥…ъћђ„ ≤ъЉ∞Јюќсє¶ƒ№Љџ÷µ„џЇѕ∆јєј±н
ЄщЊЁƒ№÷µЈ÷ќцЈљЈ®ЇЌ≤љ÷и(ЌЉ1-27)ґ‘√њ“ї÷÷„ ‘іµƒƒ№Ѕњ°Ґќп÷ љш––Ј÷ќц£ђ»°µ√“їѕµЅ–їщ±Њ эЊЁ£ђ‘ЏіЋїщі°…ѕ±а÷∆…ъћђ„ ≤ъЉ∞∆дЈюќсє¶ƒ№„џЇѕ∆јєј±н(±н1-31ЇЌ±н1-32)°£
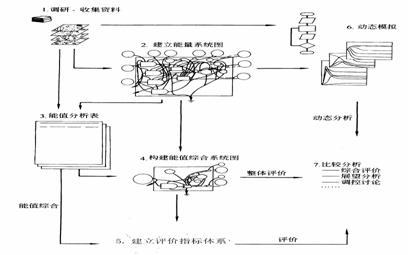
ЌЉ1-27ƒ№÷µЈ÷ќц≤љ÷и(Ј¬јґ ҐЈЉ£ђ2002;Odum,1996)
±н1-31 ƒ№÷µЈ÷ќц±н
|
„ ‘іја±р |
„ ‘іЅчґѓЅњ£®µ•ќї/ƒк£© |
ћЂ—фƒ№÷µ„™її¬ £®sej/J£© |
ћЂ—фƒ№÷µ£®sej£© |
ЇкєџЊ≠Љ√Љџ÷µ
£®EM$£© |
|
1.±ЊµЎ„ ‘і
1£©
2£©
3£©
£Ѓ£Ѓ£Ѓ
2. д»л„ ‘і
£Ѓ£Ѓ£Ѓ
3. д≥ц„ ‘і
£Ѓ£Ѓ£Ѓ |
|
|
|
|
±н1-32 ≤ЁµЎ…ъћђ„ ≤ъЉ∞∆дЈюќсє¶ƒ№Љџ÷µ„џЇѕ∆јєј±н
|
„ ‘іја±р |
„ ‘іЅчґѓЅњ |
µ•ќї |
ћЂ—фƒ№÷µ„™її¬ |
ћЂ—фƒ№÷µ£®sej£© |
ЇкєџЊ≠Љ√Љџ÷µ£®EM$£© |
|
1.Јюќсє¶ƒ№Љџ÷µ
1£© ’фЈҐ
2£© GPP
3£© NPP
1£© ћЉїг
2£© ЌЈ≈—х∆ш
3£© ≤ъ∆Ј
4£© ”йј÷
£Ѓ£Ѓ£Ѓ
2.…ъћђ„ ≤ъЉџ÷µ
1£©ЅҐµЎ∆љћ®Љџ÷µ
2£©їоµƒ…ъќпЅњ
3£©ќп÷÷їщ“т
4£©ƒаћњЉ∞”–їъ÷
£Ѓ£Ѓ£Ѓ
3.ћжіъЉџ÷µ
1£©≤њЈ÷ћжіъЉџ÷µ£®»з±н≤г…ъќпЅњћжіъЉџ÷µ£©
2£©Ќк»ЂћжіъЉџ÷µ |
|
|
|
|
|
9.3 ќэЅ÷єщј’—т≤Ё—щЈљ≤ЁµЎ…ъћђ„ ≤ъЉ∞є¶ƒ№Љџ÷µµƒƒ№÷µ∆јєј
ќ™ЅЋћљћ÷ќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥µƒЉџ÷µ£ђ—°‘сќї”ЏЄ√«шЊ≥ƒЏµƒ÷–єъњ∆—І‘ЇƒЏ√…≤Ё‘≠…ъћђѕµЌ≥ґ®ќї’Њµƒ—щµЎ„ Ѕѕќ™їщі°£ђ ‘ЌЉЌ®єэ∞Єјэ—–Њњ£ђ’є Њ≤ЁµЎ…ъћђѕµЌ≥Љџ÷µ∆јєјµƒ–¬ЈљЈ®°£
±Њ—–Њњ“‘—т≤Ё—щµЎ„ Ѕѕќ™јэљш––ЅЋ—–Њњ°£—т≤Ё—щµЎљ®”Џ1980ƒк£ђ√жїэ400m*600m°£
9.3.1 µ•ќї√жїэƒ№÷µ∆јєј
ƒЏ√…є≈ќэЅ÷єщј’µд–Ќ≤ЁµЎ—т≤Ё≤ЁµЎ…ъћђѕµЌ≥µ•ќї√жїэƒ№÷µ∆јєјљбєыЅ–”Џ±н1-33÷–£ђ≤Ґґ‘”–єЎ«эґѓЅ¶°ҐїЈЊ≥ЈюќсЉ∞…ъћђ„ ≤ъµƒЇкєџЊ≠Љ√Љџ÷µ“‘ƒ№÷µ‘™(em¥)ќ™µ•ќїљш––ЅЋєјЋг°£
|
±н1-33 |
|
|
–тЇ≈ |
ѕоƒњ |
эЊЁЉ∞ |
µ•ќї |
ƒ№÷µ„™її¬ |
ћЂ—фƒ№÷µ |
ЇкєџЊ≠Љ√Љџ÷µ |
|
|
|
|
|
(sej/µ•ќї) |
(sej/a) |
(‘™»Ћ√с±“) |
|
|
«эґѓЅ¶£Їƒ№Ѕњјі‘і |
|
|
|
|
|
|
1 |
ћЂ—фЈш…д |
4.43E+13 |
J/hm2/a |
1 |
4.43E+13 |
88.60 |
|
2 |
Јз |
3.10E+10 |
J/hm2/a |
1496 |
4.64E+13 |
92.75 |
|
3 |
”кЋЃ£Їїѓ—Іƒ№ |
1.73E+10 |
J/hm2/a |
18199 |
3.14E+14 |
629.69 |
|
4 |
”кЋЃ£Ї ∆ƒ№ |
6.49E+07 |
J/hm2/a |
27874 |
1.81E+12 |
3.62 |
|
5 |
µЎ«т—≠їЈ |
1.00E+10 |
J/hm2/a |
34377 |
6.79E+14 |
687.54 |
|
|
є¶ƒ№£Ї÷ч“™їЈЊ≥Јюќсє¶ƒ№ |
|
|
|
|
7285.94 |
|
6 |
ЋЃЈ÷’фЈҐ(”√ЋЃЅњ) |
1.98E+10 |
J/hm2/a |
18199 |
3.60E+14 |
720.68 |
|
7 |
NPP |
1.19E+11 |
J/hm2/a |
13788 |
1.04E+15 |
3281.54 |
|
8 |
GPP |
6.31E+11 |
J/hm2/a |
2602 |
1.04E+15 |
3283.72 |
|
|
љбєє£Ї÷ч“™…ъћђ„ ≤ъ |
|
|
|
|
10580.06 |
|
9 |
їо…ъќпЅњ |
1.69E+11 |
J/hm2 |
27576 |
4.16E+15 |
9320.69 |
|
10 |
≤ЁµЎЋЃЈ÷ЇђЅњ |
3.46E+10 |
J/hm2 |
18199 |
6.29E+14 |
1259.37 |
„Ґ£Ї1°Ґ±Њ±н“‘1hm2—т≤Ё—щµЎќ™јэљш––Љ∆Ћг°£1hm2≤ЁµЎ…ъћђѕµЌ≥µƒ«эґѓЅ¶°Ґ…ъћђЈюќсє¶ƒ№Љ∞∆д…ъћђ„ ≤ъµƒЇкєџЊ≠Љ√Љџ÷µµ»”ЏЄчѕоƒњµƒƒ№÷µ≥э“‘÷–єъƒ№÷µїх±“±»¬ (5X1011sej/RMB,—ѕ√ѓ≥ђ£ђ2001)£ї
2°ҐNPPЇЌGPPЉ∞їо…ъќпЅњµƒƒ№÷µ„™її¬ јі„‘±Њ—–Њњ£ђ∆дЋыµƒƒ№÷µ„™її¬ јі„‘”ЏєъЉ Ќ®”√÷Є±к£ї
і”±н1-33њ…“‘њі≥ц£ђ1hm2≤ЁµЎ…ъћђѕµЌ≥ƒкЊщћбє©µƒ÷ч“™їЈЊ≥Јюќсє¶ƒ№‘Љќ™7285.94‘™/hm2/a,£ђ∆д÷–ЋЃЈ÷’фЈҐЉџ÷µќ™720.68‘™(em ¥)/hm2/a(іЋЈюќсє¶ƒ№њ…“‘ єїЈЊ≥±£≥÷“їґ®µƒ ™ґ»£ђ¬ъ„г…ъћђѕµЌ≥ґѓќпЇЌ÷≤ќпµƒ–и«у)£ђЊї…ъќпЅњ(NPP)µƒЉџ÷µќ™3281.54‘™/hm2/a(іЋѕоЈюќсє¶ƒ№њ…“‘¬ъ„г…ъћђѕµЌ≥ЄчјаґѓќпЉ∞ќҐ…ъќпµ»µƒ–и«у)£ђ≥хЉґ…ъќп…ъ≤ъЅ¶(GPP)µƒЉџ÷µќ™3283.72‘™(іЋѕоЈюќсє¶ƒ№њ…“‘¬ъ„г…ъћђѕµЌ≥‘Ћ„™£ђќь ’CO2µ»∆шће≤Ґћеѕ÷“їѕµЅ–ѕаєЎµƒ…ъћђЈюќсє¶ƒ№)°£
1hm2≤ЁµЎ…ъћђѕµЌ≥µƒ…ъћђ„ ≤ъ‘Љќ™10580.06‘™/hm2£ђ∆д÷–їо…ъќпЅњµƒЉџ÷µ‘Љќ™9320.69‘™/hm2,∞йЋж„≈…ъћђ„ ≤ъµƒљшїѓґш–оїэµƒЋЃЈ÷µƒ…ъћђ„ ≤ъµƒЉџ÷µќ™1259.37‘™/hm2°£
9.3.2 ќэЅ÷єщј’—т≤Ё≤ЁµЎ…ъћђѕµЌ≥µƒћжіъЉџ÷µ
ќ“√«Љў…иќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥ї÷Єі∆Џќ™8ƒк£ђ≤ҐЊЁіЋ∆јєј≤ЁµЎ…ъћђѕµЌ≥µƒћжіъЉџ÷µ°£…ъћђѕµЌ≥µƒћжіъЉџ÷µV=Vs+Vc, љ÷–Vs±н Њ…ъћђѕµЌ≥Јюќсє¶ƒ№µƒћжіъЉџ÷µ£ђVcіъ±н…ъћђ„ ±ЊµƒћжіъЉџ÷µ°£…ъћђѕµЌ≥µƒЈюќсє¶ƒ№µƒћжіъЉџ÷µVs=1/2*T*(GPP*TrGPP /P+VNPP+VЋЃ)£ђ љ÷–GPPќ™…ъћђѕµЌ≥µƒ≥хЉґ…ъ≤ъЅ¶£ђґшTrGPP ќ™GPPµƒƒ№÷µ„™її¬ , Tќ™ћжіъ ±Љдїтї÷Єі ±Љд(replacement time), Pќ™ƒ№÷µ/їх±“±»¬ £ђVNPPЇЌVЋЃЈ÷±ріъ±нЊї…ъќпЅњЇЌ≤ЁµЎЋЃЈ÷÷ьЅњµƒЉџ÷µ°£ЄщЊЁ…ѕ цЈ÷ќц£ђ…ъћђѕµЌ≥µƒћжіъЉџ÷µV= 1/2*(GPP*TrGPP*T/P + T VNPP+ T VЋЃ)+Vc£ђЉ∆ЋгљбєыЉы±н1-34°£
±н1-34 1hm2ќэЅ÷єщј’—т≤Ё≤ЁµЎ…ъћђѕµЌ≥µƒћжіъЉџ÷µ∆јєј
|
ѕо ƒњ |
Љ∆ЋгєЂ љ |
ЇкєџЊ≠Љ√Љџ÷µ(em$) |
±Є„Ґ |
|
1.їЈЊ≥Јюќсє¶ƒ№ћжіъЉџ÷µ(Vs) |
Vs=1/2*T*(GPP*TrGPP /P+VNPP+VЋЃ) |
29143.76 |
GPPЇЌTrGPP эЊЁјі‘і”Џ±н5-2£ђTќ™8ƒк°£ |
|
2.…ъћђ„ ±Њ(Vc) |
Vc=Vc1+Vc2+°≠+Vcn |
10580.06 |
Vc1,Vc2, °≠,Vcnіъ±н≤їЌђја–Ќ…ъћђ„йЈ÷µƒ…ъћђ„ ±Њ°£ |
|
„№ћжіъЉџ÷µ |
V=Vs+Vc |
|
|
9.3.3 ќэЅ÷єщј’—т≤Ё—щµЎ…ъћђѕµЌ≥Љџ÷µµƒ„џЇѕєјЋг
‘ЏЉ∆Ћг≤ЁµЎ…ъћђѕµЌ≥µƒћжіъЉџ÷µ ±£ђќ“√«Љў…и≤ЁµЎ…ъћђѕµЌ≥Ќк»Ђѕы І°£ћжіъЉџ÷µ «≤ЁµЎ…ъћђѕµЌ≥ЈюќсЉџ÷µ”л„‘»ї„ ±ЊЉџ÷µµƒ„№ЇЌ°£ЄщЊЁќэЅ÷єщј’√Ћµƒ µЉ £ђ1hm2—т≤Ё—щµЎ…ъћђѕµЌ≥ћжіъЉџ÷µ‘Љќ™39724‘™£їі”24hm2—щµЎµƒ«йњцјіњі£ђ∆д„№ћжіъЉџ÷µ‘Љќ™953371‘™(±н1-35)°£
±н1-35 ќэЅ÷єщј’24hm2њ…јы”√≤Ё≥°…ъћђѕµЌ≥Љџ÷µ„џЇѕ∆јєјљбєы
|
ѕоƒњ |
√жїэ(hm2) |
ЇкєџЊ≠Љ√Љџ÷µ(‘™) |
µ•ќї |
|
“ї°ҐїЈЊ≥Јюќсє¶ƒ№ |
24 |
174862.56 |
em¥/a |
|
ґю°Ґ…ъћђ„ ±Њ |
24 |
253921.44 |
em¥ |
|
»э°Ґ„№ћжіъЉџ÷µ |
24 |
953371.68 |
em¥ |
ЄщЊЁ“‘…ѕµд–Ќ…ъћђѕµЌ≥µƒ—–Њњљбєы£ђќ“√«њ…“‘ґ‘ќэЅ÷єщј’24hm2—т≤Ё—щµЎ≤Ё≥°µƒЉџ÷µ„ч≥х≤љєјЋг°£
9.4 –°љб
9.4.1 ƒ№÷µ”л≤ЁµЎ…ъћђѕµЌ≥µƒЇкєџЊ≠Љ√Љџ÷µ
‘Ћ”√ƒ№÷µјн¬џґ‘…ъћђѕµЌ≥µƒЉџ÷µљш––∆јєј£ђ”√ƒ№÷µ‘™(em$їтem¥)ґ‘…ъћђѕµЌ≥µƒљбєєЉ∞∆дїЈЊ≥Јюќсє¶ƒ№µƒЉџ÷µљш––∆јєј£ђњ…“‘ єЉџ÷µ∆јєјљ®ЅҐ‘Џњ∆—ІњЌєџµƒїщі°÷Ѓ…ѕ°£±Њ—–Њњљбєы±н√ч£ђ‘ЏќэЅ÷єщј’£ђ1hm2≤ЁµЎ…ъћђѕµЌ≥∆љЊщ√њƒкґ‘«ш”тЊ≠Љ√є±ѕ„µƒЇкєџЊ≠Љ√Љџ÷µ‘Љќ™7285.94‘™£ђґш1hm2≤ЁµЎ…ъћђѕµЌ≥÷ч“™…ъћђ„ ≤ъµƒЇкєџЊ≠Љ√Љџ÷µ‘Љќ™10580.06‘™°£і”24hm2—щµЎµƒ«йњцјіњі£ђ…ъћђѕµЌ≥Јюќсє¶ƒ№µƒЉџ÷µ‘Љќ™174863‘™/a£ђ…ъћђ„ ±ЊµƒЉџ÷µ‘Љќ™253941‘™(±н5-4)°£»зєыƒ≥“їЊ≠Љ√њ™ЈҐ––ќ™µЉ÷¬’в“ї«ш”т≤ЁµЎ…ъћђѕµЌ≥Ћр І£ђќ“√«њ…“‘”√…ѕ ц—–ЊњљбєыєјЋгїЈЊ≥Јюќсє¶ƒ№Љ∞…ъћђ„ ±ЊµƒЋр І«йњц£ђ“≤њ…“‘ЄщЊЁ…ъќпЅњµƒЉх…ўЉ∞GPPµƒ±дґѓ«йњцґ‘…ъћђѕµЌ≥µƒЈюќсє¶ƒ№Љ∞…ъћђ„ ±Њ±дґѓ«йњцљш––∆јєј°£Ќт“їƒ≥“їµЎ«ш≤ЁµЎ…ъћђѕµЌ≥ЌЋїѓ£ђќ“√«њ…“‘ЄщЊЁ≤ЁµЎ…ъћђѕµЌ≥є¶ƒ№ЌЋїѓµƒ≥ћґ»єјЋг≥ц«ш”т…ъћђѕµЌ≥Ћр Іµƒ«йњц°£
9.4.2 ƒ№÷µ…ъћђЉџ÷µ∆јєјЈљЈ®µƒњ∆—І–‘Љ∞”¶”√«∞Њ∞
”ліЂЌ≥µƒЊ≠Љ√—ІЈљЈ®ЇЌ÷ІЄґ“в‘ЄЈљ љѕа±»£ђ‘Ћ”√ƒ№÷µјн¬џљш––…ъћђѕµЌ≥µƒЉџ÷µ∆јєјµƒЈљЈ®љѕќ™њЌєџєЂ’э£ђњЋЈюЅЋіЂЌ≥ЈљЈ®µƒ÷чєџ…Ђ≤ £ђЊя”–—ѕ√№µƒњ∆—Іїщі°ЇЌљѕќ™њЌєџµƒЉџ÷µ≥яґ»£ђ“тґшЊя”–єгЈЇµƒ”¶”√«∞Њ∞°£
9.5 ќэЅ÷єщј’≤ЁµЎ÷≤±ї≤г…ъќпЅњ„ ≤ъЉ∞∆дЈюќсє¶ƒ№Љџ÷µґѓћђƒ£ƒв
9.5.1 ÷ч“™…ъћђ„ ≤ъЉ∞÷Ў“™Јюќсє¶ƒ№Љџ÷µ∆јєјљбєы
Ќ®єэґ‘ќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥ЉЄѕо÷ч“™…ъћђ„ ≤ъµƒЉџ÷µљш––∆јєј£ђљбєы±н√ч£®±н1-36£©£Ї…ъќпЅњ…ъћђ„ ≤ъ(їо…ъќпЅњ)µƒЉџ÷µќ™1651“Џ‘™»Ћ√с±“£ђ÷ч“™…ъћђѕµЌ≥Јюќсє¶ƒ№µƒЉџ÷µјџЉ∆ќ™1651“Џ‘™£ђ÷ч“™ћжіъЉџ÷µ8255“Џ‘™°£ћжіъЉџ÷µ∆јєјљбєы±н√ч£ђ»зєыќэЅ÷Єсј’≤ЁµЎ…ъћђѕµЌ≥‘вµљ—ѕ÷Ў∆∆їµ£ђ“™ї÷Єі…ѕ цЉЄѕо÷ч“™є¶ƒ№÷Ѕ…ў–и“™Ќґ„ 8255“Џ‘™°£
”…”Џ –≥° ІЅйЇЌЉџЄсЄ№ЄЋµƒ≈§«ъ£ђ–нґа…ъћђЊ≠Љ√Љџ÷µќіƒ№‘ЏіЂЌ≥µƒЈљЈ®÷–µ√“‘ћеѕ÷£ђЉџ÷µ±їµЌєјЅЋ£ђљцЉЄѕо÷ч“™µƒ…ъћђѕµЌ≥Јюќсє¶ƒ№Љџ÷µ£®1651“Џ‘™/ƒк£©ЊЌіуіу≥ђєэЅЋќэЅ÷єщј’≈©Ѕ÷ƒЅ”ж“µ≤ъ÷µµƒ„№ЇЌ£®33.97“Џ‘™£©°£ќ™ЅЋ√÷≤є –≥°Њ≠Љ√ЇЌЉџЄсћеѕµµƒ»±ѕЁ£ђєъЉ“”¶÷∆ґ®≥ц“їґ®µƒ≤є≥•їъ÷∆£ђґ‘’в“ї«ш”т≤ЁµЎ…ъћђѕµЌ≥µƒљ®…и±£ї§”и“‘÷І≥÷°£
±н1-36°°÷ч“™…ъћђ„ ≤ъЉ∞Јюќсє¶ƒ№Љџ÷µ∆јєј±н
|
Љџ÷µја–Ќ |
Љ∆ЋгєЂ љ |
Љџ÷µ£®“Џ‘™£© |
|
1.…ъќпЅњ…ъћђ„ ≤ъ |
VQ£љ923.5Q£®‘™£© |
1651 |
|
2.Јюќсє¶ƒ№Љџ÷µ |
Es=Fc+Fo+So |
1651 |
|
2.1єћґ®CO2Љџ÷µ |
Fc=1.63NPP*P=1.63*NPP*250(‘™/t) =407.5NPP(‘™) |
371 |
|
2.2 ЌЈ≈O2Љџ÷µ |
Fo=1.19NPP*Po=1.19*500NPP=595NPP(‘™) |
541 |
|
2.3ћбє©”–їъќпЉџ÷µ |
So£љ923.5(J2+J4)£®‘™£© |
739 |
|
3.ћжіъЉџ÷µ |
Vs= 1/2(Fc+Fo+So)*8+ VQ =4(Fc+Fo+So)+ VQ |
8255 |
9.5.2 ќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥…ъќпЅњ„ ≤ъЉ∞÷ч“™Јюќсє¶ƒ№Љџ÷µґѓћђ±дїѓ
ЄщЊЁ…ъќпЅњЉ∞÷ч“™є¶ƒ№÷µµƒƒ£ƒвљбєы£ђљбЇѕ…ъћђ„ ≤ъЉ∞Јюќсє¶ƒ№Љ∆ЋгЈљ≥ћ£ђќ“√«њ…“‘µ√µљ…ъќпЅњ…ъћђ„ ≤ъЉ∞Јюќсє¶ƒ№ґѓћђ±дїѓµƒƒ£ƒвљбєы£®ЌЉ1-28£©°£і”ЌЉ

ЌЉ1-28 ќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥÷ч“™…ъћђє¶ƒ№÷µ±дїѓґѓћђƒ£ƒвљбєы
÷–њ…“‘њі≥ц£ђ1992ƒкЄ√«ш≤ЁµЎ…ъћђѕµЌ≥”…”Џ‘Ў–уЅњ≤Ґќіƒ№іпµљ„оіу÷µ£ђґ‘≤ЁµЎµƒ…ъќпЅњѕыЇƒЅњµЌ”Џ…ъ≥§Ѕњ£ђ≤ЁµЎ‘Ў–уЅњ…–”–“їґ®µƒ‘ц≥§«±Ѕ¶°£’в÷÷«йњц”л1992ƒк“‘јі≤Ё–у эЅњµƒ‘ц≥§µƒ ¬ µЋщ÷§√ч£ђЋµ√чіЋƒ£ƒвљбєы”л ¬ µѕаЈы°£
ƒ£ƒвљбєы±н√ч£ђі”„№ће«йњцњі£ђ„‘і”1992ƒк“‘јі£ђќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥÷≤±ї≤г…ъќпЅњ…ъћђ„ ≤ъЉ∞÷ч“™Јюќсє¶ƒ№µƒЉџ÷µ≥ ѕ÷≥ц‘ц≥§ћђ ∆£ђµљ2004ƒк£ђЄчѕоЈюќсє¶ƒ№÷µ≤Ґќііпµљґ•ЉЂ„іћђ£ђ‘ц≥§…–”–“їґ®µƒ«±Ѕ¶°£2004ƒк£ђ…ъќпЅњ…ъћђ„ ≤ъµƒЉџ÷µќ™2421“Џ‘™£ђ÷ч“™Јюќсє¶ƒ№Љџ÷µ1902“Џ‘™/ƒк£®∆д÷–£ђєћґ®CO2Љџ÷µ407“Џ‘™£ђ ЌЈ≈O2Љџ÷µ595“Џ‘™£ђћбє©”–їъќпЉџ÷µ900“Џ‘™£©£ђ…ъћђѕµЌ≥µƒћжіъЉџ÷µіпµљ10031“Џ‘™°£ЄщЊЁƒ£ƒв‘§≤в£ђ2031ƒк£ђќэЅ÷єщј’≤ЁµЎ…ъћђѕµЌ≥µƒ÷ч“™…ъћђЊ≠Љ√Љџ÷µљЂіпµљ£Ї…ъћђ„ ≤ъ(…ъќпЅњ)µƒЇкєџЊ≠Љ√Љџ÷µќ™2585“Џ‘™£ђ÷ч“™Јюќсє¶ƒ№µƒЉџ÷µќ™1928“Џ‘™/a£ђƒ« ±£ђ…ъћђѕµЌ≥µƒћжіъЉџ÷µќ™10296“Џ‘™°£
…ѕ“ї“≥ [1] [2] [3] [4] [5] [6]
|